ТЕМА: «Физиология центральной нервной системы и
высшей нервной деятельности»
Тирасполь
УДК 572.788:612.8(07)(075.8)
ББК Е991р3я73+Р252.7р3я73
Л 47
Аннотация:
Данные методические указания позволяют ознакомиться с клинически важными рефлексами человека и механизмами их возникновения, научиться вырабатывать условные рефлексы на индифферентный раздражитель. Ознакомиться с психофизиологическим методом исследования силовых показателей в деятельности ЦНС, а также с некоторыми приемами и методами исследования памяти у человека, овладеть методами исследования некоторых высших психических функций человека.
Предложенные методические указания позволяют осуществить практическое закрепление полученных теоретических знаний.
Составители:
А.И. Леорда, доцент, к.б.н., зав. кафедрой физиологии и фармакологии медицинского факультета ПГУ им. Т.Г. Шевченко
Л.Д.Стоянова, преподаватель кафедры физиологии и фармакологии медицинского факультета ПГУ им. Т.Г. Шевченко
Рецензенты:
Павалюк П.П.,заведующий лабораторией Института физиологии и санокреатологии АНМ,
к.б.н., старший научный сотрудник.
Г.П. Пальцева, ст.преподаватель кафедры физиологии и фармакологии медицинского факультета ПГУ им. Т.Г. Шевченко
Утверждено Научно-методическим Советом
ПГУ им. Т.Г. Шевченко
Протокол № от «____» ___________2009г.
© А.И. Леорда Л.Д. Стоянова, 2009
ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
ТЕМА 1: МЕТОДЫ ИССЛЕДОВАНИЯ ФУНКЦИИ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ.
НЕОБХОДИМО ЗНАТЬ:
- Методы изучения функции ЦНС.
- Функции спинного мозга. Основные проводящие пути спинного мозга. Характеристика спинальных животных, спинальный шок. Роль спинного мозга в регуляции вегетативной функции.
- Роль спинного мозга в регуляции деятельности опорно-двигательного аппарата и мышечного тонуса. Клинически важные спинальные рефлексы.
- Продолговатый мозг и мост, их участие в регуляции вегетативных функций организма. Важнейшие защитные рефлексы.
- Децеребрационная ригидность и механизм ее возникновения, роль продолговатого мозга в регуляции мышечного тонуса.
- Физиология среднего мозга, его рефлекторная деятельность и участие в процессах регуляции функций организма. Статические и статокинетические рефлексы. Опыты Магнуса.
Функции и роли вожатого
... и даст инструкции. Способность ясно видеть перспективы, рационально использовать ресурсы Слишком жесткая регуляция, мало внимания к человеческим потребностям Нерешительность, неясная политика 5. Координатор- От координатора ...
КЛЮЧЕВЫЕ ПОНЯТИЯ И ОПРЕДЕЛЕНИЯ
Функции ЦНС изучают с помощью классических методов: удаление (экстирпация), разрушение изучаемого нервного образования, кратковременное блокирование передачи возбуждения химическим веществом, денервация органа, электроэцефалография, электронейронография, нейро- и гистохимия, стереотаксическая операция и др..
Спинальный шок – это временное угнетение функционального тонуса нервных центров, расположенных ниже области перерезки (повреждения) спинного мозга, вследствие нарушения его связей с вышележащими центрами головного мозга
Децеребрационная ригидность – патологическое состояние, возникающее при перерезке на уровне переднего края задних холмов четверохолмия и отделяющее красное ядро от вестибулярного ядра Дейтерса. Это состояние характеризуется резким повышением тонуса мышц-разгибателей, конечностей, спины и хвоста у животных.
Опыты Магнуса. Шейные тонические рефлексы запускаются при возбуждении проприорецепторов мышц шеи. Наблюдаются при разрушении вестибулярного аппарата. Запрокидывание головы кошки назад вызывает повышение тонуса мышц-разгибателей передних конечностей и расслабление мышц-экстензоров задних конечностей. При наклоне головы вниз возникает обратный эффект.
Статические рефлексы связаны с возбуждением рецепторов лабиринта и обеспечивают поддержание позы и равновесия тела при статических положениях в пространстве.
Статокинетические рефлексы направлены на поддержание позы при изменении скорости движения, возникают в результате возбуждения рецепторов полукружных каналов.
НЕОБХОДИМО УМЕТЬ:
- Приобрести навыки исследования сухожильных рефлексов у человека.
- Доказать рефлекторный механизм мышечного тонуса.
3. Установить значение рецепторов вестибулярного аппарата в регуляции мышечного тонуса.
РАБОТА №1 Рефлексы человека, имеющие клиническое значение.
Цель работы: ознакомиться с клинически важными рефлексами человека и механизмами их возникновения. Приобрести навыки исследования сухожильных рефлексов у человека.
Необходимо для работы: испытуемый, неврологический молоточек.
Проведение работы:
Коленный рефлекс
- Испытуемый сидит, положив ногу на ногу. Мышцы исследуемой конечности должны быть расслаблены. Произвести отрывистые удары молоточком по сухожилию ниже коленной чашечки. Отметить сокращение четырехглавой мышцы берда, которые вызывают разгибание голени.
- Сравнить рефлексы обеих конечностей. Если коленный рефлекс ослабленный, то испытуемый должен, прочно сцепив пальцы обеих рук, сильно растянуть их в стороны. При этом коленный рефлекс значительно усилится (феномен Ендрашика) – снимаются тормозные влияния коры на двигательные центры спинного мозга.
Ахиллов рефлекс
- Испытуемый встает коленями на стул, держась руками за его спинку. Произвести отрывистые удары по ахилловому сухожилию и отметить рефлекторные разгибательные движения стопы, наступающие вследствие сокращения трехглавой мышцы голени.
- Сравнить рефлексы на обеих ногах.
В протокол зарисовать и описать рефлекторные дуги проприорецептивных рефлексов (коленного и ахиллового), отметив их особенности. Указать, в каких сегментах спинного мозга расположены центры этих рефлексов.
1. Рефлекс Россолимо-Вендеровича
... – разгибаются конечности на данной стороне, а на другой – сгибаются. угасает в 2-3 месяца. 14. Лабиринтный тонический рефлекс – в положении на спине повышен тонус в разгибателях, ... НЕВРОЛОГИЯ Симптомы параличей Периферический паралич: 1. Арефлексия. 2. Мышечная атония. 3. Атрофия мышц. 4. Реакция перерождения при электрофизиологическом исследовании. 5. Фибриллярные или фасцикулярные подергивания. ...
РАБОТА №2. Рефлекторный тонус скелетных мышц (опыт Бронджиста).
Цель работы: в условиях эксперимента доказать рефлекторный механизм мышечного тонуса.
Необходимо для работы: лягушка, штатив с крючком для ее фиксации, препаровальный набор.
Проведение работы:
- Приготовить бульбарную лягушку.
2. Сбоку от таза сделать небольшой разрез кожи и мышц (около 1 см).
Через разрез найти поясничное сплетение. Провести под сплетение нитку. Подвесить лягушку за нижнюю челюсть. Внимательно рассмотреть положение нижних конечностей: равенство углов, образуемых голенью и бедром, а также голенью и стопой.
3. Туго перевязать и перерезать поясничное сплетение.
4. Через несколько минут сравнить длину обоих лапок, обратив внимание на увеличение углов между частями на оперированной стороне. Оперированная лапка слегка вытягивается в результате выключения тонического рефлекса растяжения.
5. В протоколе описать и зарисовать положение лапок лягушки до и после односторонней денервации. Объяснить причину тонического сокращения мышц лапки с сохраненной иннервацией.
6. Опыт можно провести и в более упрощенном варианте. Для этого у спинальной лягушки на одной лапке на дорзальной стороне бедра разрезать кожу, раздвинуть мышцы бедра, обнажить седалищный нерв и туго перевязать его при помощи лигатуры или перерезать ножницами. Произвести те же наблюдения, что и в предыдущих пунктах работы.
РАБОТА №3. Влияние лабиринтов на тонус мускулатуры
Цель работы: установить значение рецепторов вестибулярного аппарата в регуляции мышечного тонуса.
Необходимо для работы: лягушка, препаровальная игла, марлевая салфетка, таз для воды или аквариум.
Проведение работы:
1.Наблюдать в течение 5-10 мин. за нормальным положением тела лягушки (прыжки, плавательные движения).
2.Обернуть лягушку марлей, открыть рот. Нижнюю челюсть оттянуть вниз. Ножницами или скальпелем сделать крестообразный разрез слизистой оболочки дна полости рта вдоль и поперек основной кости верхней челюсти. Раздвинуть края раны и обнажить основную кость. Справа и слева сзади от глаз на поверхности основной кости найти белые бугорки. В один из них ввести препаровальную иглу и вращательными движениями разрушить лабиринт. Вытекающую кровь удалить марлевым тампоном.
3.Освободить лягушку и посадить ее на стол. Наблюдать за позой, прыжками. Обратить внимание, что при прыжках лягушка падает на бок или спину.
4.Поместить лягушку в таз или аквариум с водой. Наблюдать за плавательными движениями. Лягушка должна двигаться по кругу и переворачиваться вокруг продольной оси своего тела.
5.В протоколе кратко описать ход работы и результаты наблюдений. Объяснить механизм, лежащий в основе нарушения двигательных функций в связи с перераспределением мышечного тонуса. Сделать выводы о роли рецепторов вестибулярного аппарата в поддержании тонуса скелетной мускулатуры.
6.Лягушку в конце опыта обездвижить.
ТЕМА 2: ЧАСТНАЯ ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ.
НЕОБХОДИМО ЗНАТЬ:
1. Ретикулярная формация ствола мозга, особенности нейронов ретикулярной формации. Нисходящие и восходящие влияния ретикулярной формации. Участие ретикулярной формации в формировании целостной деятельности организма.
2. Таламус. Функциональная характеристика и особенности ядерных групп таламуса.
3. Гипоталамус. Характеристика основных ядерных групп. Участие гипоталамуса в иннервации рефлекторных и гормональных механизмов регуляции функций и в формировании мотиваций и эмоций.
4. Физиология мозжечка, его влияние на моторные вегетативные функции организма. Основные симптомы удаления или частичного поражения мозжечка.
5. Лимбическая система мозга. Ее роль в формировании биологических мотиваций и эмоций.
6. Базальные ганглии, их роль в регуляции мышечного тонуса и сложных двигательных актов. Основные симптомы поражения базальных ганглиев.
7. Методы изучения деятельности коры больших полушарий. Метод электроэнцефалографии (ЭЭГ), основные ее ритмы.
КЛЮЧЕВЫЕ ПОНЯТИЯ И ОПРЕДЕЛЕНИЯ
Ретикулярная формация ствола мозга представлена сетью нейронов с многочисленными диффузными связями между собой и со всеми структурами ЦНС. Основной функцией является регуляция уровня активности коры большого мозга, мозжечка, таламуса, спинного мозга. Может избирательно оказывать активирующее или тормозящее влияние на разные формы поведения, сенсорные, моторные, висцеральные системы мозга.
Таламус – структура, в которой происходит обработка и интеграция всех сигналов, идущих в кору большого мозга от всех образований ЦНС. Ядра таламуса делятся на специфические, неспецифические и ассоциативные. Коленчатые тела таламуса являются подкорковыми центрами зрения и слуха, узел уздечки и переднее зрительное ядро анализируют обонятельные сигналы. Таким образом, зрительный бугор – подкорковая «станция» для всех видов чувствительности.
Гипоталамус – филогенетически старый отдел промежуточного мозга, играющий важную роль в поддержании гомеостаза и обеспечивающий интеграцию функций вегетативной, эндокринной и соматической систем. В нем выделяют 50 пар ядер, топографически объединенные в пять групп: преоптическая, передняя, средняя, наружная, задняя.
Лимбическая система — это функциональное объединение структур мозга, участвующих в организации эмоционально-мотивационного поведения (пищевой, половой, оборонительный инстинкты).
С ее помощью формируются нейронные механизмы памяти, психические и вегетативные функции.
Базальные ганглии — структуры располагающиеся преимущественно у основания лобных долей, к ним относятся хвостатое ядро, ограда, бледный шар, скорлупа, полосатое тело.
НЕОБХОДИМО УМЕТЬ:
1. Установить роль различных отделов ЦНС в регуляции мышечного тонуса и фазной активности.
РАБОТА №4. Роль различных отделов ЦНС в формировании мышечного тонуса и фазных движений
Цель работы: установить роль различных отделов ЦНС в регуляции мышечного тонуса и фазной активности.
Необходимо для работы: лягушка, препаровальный набор, марлевая салфетка.
Проведение работы:
- Лягушку завернуть в марлевую салфетку так, чтобы голова ее оставалась открытой.
2. Обнажить головной мозг. Для этого позади ноздрей ножницами сделать поперечный разрез кожи, от краев которого провести два разреза до туловища и удалить образовавшийся кожный лоскут. Маленькими ножницами сделать поперечный разрез кости позади ноздрей и, прижимая бранши ножниц к черепу, осторожно срезать черепные кости с двух сторон до туловища. Обнажившийся головной мозг подсушить ватным тампоном.
3. Выключение различных отделов мозга (большие полушария, промежуточный, средний и продолговатый мозг) провести путем поперечных перерезок и последующей экстирпации.
4. По мере экстирпации различных отделов головного мозга пронаблюдать за сохранением позы, характерной для сидящего животного и проверить наличие рефлекса переворачивания.
5. Результаты опыта внести в таблицу.
| Лягушка | Наблюдаемые явления | ||
| Вид мышечного тонуса | Наличие позы сидящего животного | Рефлекс переворачивания | |
| Нормальная Без полушарий и таламической области Бульбарная Спинальная | Нормальный пластический контактильный простейший | + + + — | + + — — |
Выводы: Нормальная поза, характерная для сидящего животного, отсутствует у бульбарной и спинальной лягушек. У спинальной лягушки нет рефлексов переворачивания. Таким образом, нормальная двигательная активность и мышечный тонус регулируются при участии всех отделов центральной нервной системы.
ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ ПСИХИЧЕСКОЙ ДЕЯТЕЛЬНОСТИ
ТЕМА 3: МЕТОДЫ ИЗУЧЕНИЯ ФУНКЦИЙ КОРЫ БОЛЬШИХ ПОЛУШАРИЙ. УСЛОВНЫЙ РЕФЛЕКС, ЕГО НЕЙРОФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ. ТОРМОЖЕНИЕ УСЛОВНО-РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ.
НЕОБХОДИМО ЗНАТЬ:
1. Условный рефлекс как метод изучения высшей нервной деятельности. Правила образования условного рефлекса.
2. Механизм образования временных связей (И.П. Павлов, современные представления).
3. Стадии выработки условных рефлексов. Классификация условных рефлексов.
4. Явление торможения в высшей нервной деятельности. Виды коркового торможения.
5. Современные представления о механизмах кратковременной и долговременной памяти.
6. Физиологические механизмы сна. Фазы сна. Теории сна. Физиологическая роль сновидений.
7. Аналитико-синтетическая деятельность коры больших полушарий. Корковый динамический стереотип, его физиологическая сущность, значение для приобретения трудовых навыков и обучения.
8. Мотивации. Периферические и центральные механизмы формирования мотиваций. Классификация мотиваций.
КЛЮЧЕВЫЕ ПОНЯТИЯ И ОПРЕДЕЛЕНИЯ:
Различные формы поведения принято разделять на врожденные (безусловные) и приобретенные (условные) в процессе индивидуального развития. Наследственно закрепленные рефлексы лежат в основе приспособительных поведенческих актов, проявляющихся без предварительного обучения.
Элементарные безусловные рефлексы являются простыми ответными реакциями местного значения, реализуемыми в соответствии с жесткой детерминированной программой своих сегментарных центров. Координационныебезусловные рефлексы осуществляются на сегментарном уровне, но допускают координацию на основе отрицательных и положительных обратных связей (работа мышц-антагонистов, формирующих сложные двигательные акты).
Интегративные безусловные рефлексы – это синтез координированных двигательных актов с их вегетативным обеспечением в комплексные реакции определенного биологического значения.
Генетически детерминированные формы поведения, отражающие накопленный в генофонде видовой опыт предшествующих поколений, оказываются недостаточными, чтобы обеспечить активное существование особи в вероятно изменчивой среде.
В основе индивидуального опыта (приобретенные формы поведения), который приобретается различными путями, лежит общая способность живых организмов к обучению, которая возможна благодаря такому универсальному свойству материи и живых организмов, как память.
Условные рефлексы –это индивидуально приобретенные системные приспособленные реакции животных и человека, возникающие на основе образования в центральной нервной системе временной нервной связи между условными (сигнальными) раздражителями и безусловно-рефлекторными актами.
Мотивация —субъективно окрашенное состояние, возникающее на основе активации мозговых структур, побуждающих высших животных и человека совершать действия, направленные на удовлетворение своих биологических потребностей.
Биологические мотивацииформируются на основании отклонений тех или иных показателей внутренней среды организма от уровня, обеспечивающего оптимальное течение обмена веществ. Биологические мотивации направлены на удовлетворение ведущих биологических потребностей – пищевых, половых, оборонительных, а также сна и бодрствования.
Социальные мотивации(высшие мотивации) возникают под влияний социальных факторов. Формируются они в процессе общественного воспитания, которое у человека оказывает значительное влияние на биологические мотивации и делает их в какой-то мере социально детерминированными.
.
НЕОБХОДИМО УМЕТЬ:
1. Научиться вырабатывать условные рефлексы на индифферентный раздражитель.
2. Ознакомить студентов с некоторыми приемами и методами исследования памяти у человека.
РАБОТА №5. Образование условного мигательного рефлекса на звонок у человека
Цель работы: научиться вырабатывать условные рефлексы на индифферентный раздражитель и наблюдать за воспроизведением условного рефлекса при замене индифферентного физического сигнала словесным.
Необходимо для работы: установка для выработки условного мигательного рефлекса у человека.
Проведение работы:
1. Собрать установку для выработки условного мигательного рефлекса у человека.
2. Установить стеклянный наконечник, соединенный с резиновой грушей так, чтобы при нажатии ее струя воздуха попала на веко наружного угла глаза.
3. Включить электрический звонок в сеть.
4. При поддувании воздуха возникает безусловный защитный мигательный рефлекс.
5. Для выработки условного рефлекса включить звонок на пять секунд и во время его звучания несколько раз поддувать воздух (время отставания безусловного раздражителя –
1 -2 секунды).
6. Через несколько сочетаний двух раздражителей у испытуемого вырабатывается условный рефлекс – изолированное включение звонка сопровождается миганием.
При образовании прочного условного мигательного рефлекса включение звонка можно заменить словесным раздражением «включаю звонок» и пронаблюдать за условной реакцией на словесный сигнал.
7. При выработке условного мигательного рефлекса на звонок необходимо установить, через сколько сочетаний образуется условный рефлекс у лиц различного темперамента.
8. В протоколах зарисовать установку для выработки условного мигательного рефлекса у человека, описать технику выработки условного рефлекса. Внести полученные результаты (через сколько сочетаний образовался условный рефлекс у разных лиц) и объяснить, почему при замене звонка словесным сигналом «включаю звонок» рефлекс возникает.
РАБОТА №6. Исследование у человека непосредственной и оперативной памяти
Цель работы: ознакомить студентов с некоторыми приемами методами исследования памяти у человека.
Проведение работы:
Тестированию могут подвергаться студенты при их добровольном согласии. Тестирование проводится только преподавателем. Результаты тестирования не регистрируются и не оглашаются.
Непосредственная память.Принцип тестирования состоит в том, что испытуемому предлагают внимательно прослушать и запомнить ряд слов либо цифр, а затем по памяти записать их.
В данном тесте исследуется возможность образования и воспроизведения сразу нескольких временных связей, замыкающихся на звуковые раздражители второй сигнальной системы.
Оперативная память. Под этим термином понимается избирательное кратковременное сохранение информации, необходимое для выполнения текущей деятельности и достижения ее целей.
Опосредованная памятьобычно исследуется с помощью пиктограмм. Испытуемый берет лист бумаги и карандаш. Затем дается инструкция: «Запомните название слова, предмета, явления. Для облегчения запоминания сделайте на бумаге зарисовки к каждому слову. Писать само слово или обозначать его буквами или цифрами нельзя».
Лист с пиктограммами сдается преподавателю и через час возвращается студентам. По рисункам студенты должны вспомнить слова и написать их под рисунками.
ТЕМА 4: ТИПЫ ВЫСШЕЙ НЕРВНОЙ ДЯТЕЛЬНОСТИ. УЧЕНИЕ И.П. ПАВЛОВА О ПЕРВОЙ И ВТОРОЙ СИГНАЛЬНЫХ СИСТЕМАХ. ВЫСШИЕ ПСИХИЧЕСКИЕ ФУНКЦИИ.
НЕОБХОДИМО ЗНАТЬ:
1. Учение И.П. Павлова о типах высшей нервной деятельности (ВНД), их классификация и характеристика.
2. Невроз (срыв высшей нервной деятельности).
Фазы невроза, механизм возникновения. Экспериментальные методы получения невроза.
3. Биологическая роль эмоций. Теории эмоций. Вегетативные и соматические компоненты эмоций. Эмоциональный стресс и его значение в формировании психо-соматических заболеваний.
4. Учение И.П. Павлова о 1 и 2 сигнальных системах. Взаимоотношение 1 и 2 сигнальных систем. Типы ВНД, свойственные только человеку.
5. Архитектура целостного поведенческого акта с точки зрения теории функциональной системы П.К. Анохина.
6. Понятие о высших психических функциях человека (ощущение, восприятие, представление, память, эмоции, мышление, речь, сознание).
7. Физиологические основы трудовой деятельности человека. Механизм адаптации организма к статическим и динамическим физическим нагрузкам.
8. Особенности умственного труда. Умственное утомление. Нервные, вегетативные и эндокринные изменения при умственном труде.
9. Адаптация организма к физическим, биологическим и социальным факторам. Виды адаптации. Значение нервных, эндокринных и генетических механизмов. Биологические ритмы, их классификация.
10. Стресс. Механизм общего адаптационного синдрома (Селье).
Особенности адаптации человека к действию экстремальных факторов.
КЛЮЧЕВЫЕ ПОНЯТИЯ И ОПРЕДЕЛЕНИЯ:
Типы ВНД: сангвинический, флегматический, холерический, меланхолический. В основе этой классификации лежат три характеристики: сила, подвижность и уравновешенность.
Невроз – патологическое состояние, возникающее в результате срыва ВНД.
Эмоция – специфическое состояние психической сферы, обусловленное потребностями организма и уровнем возможного их удовлетворения. Эмоция этореакция в виде субъективно окрашенного переживания индивида, отражающая значимость для него воздействующего раздражителя или результата собственной деятельности (удовлетворение или неудовлетворение).
Эмоциональное возбуждение формируется первично в подкорковых структурах лимбической системы. Оно оказывает восходящее влияние на расположенные выше структуры мозга, а также нейрогуморальным путем — непрерывные, генерализованные нисходящие влияния на соматовисцеральные функции
Развитие представлений о механизмах формирования целенаправленной деятельности нашло свое отражение в теории функциональных систем (П.К. Анохин).
По концепции Анохина, начальная стадия поведения осуществляется на уровне мозговых структур, формирующих афферентный синтез. Основными его компонентами являются мотивационное возбуждение, обстановочная афферентация, аппарат памяти (прошлый опыт) и пусковая афферентация (действие раздражителей, запускающих цепь рефлекторных реакций).
Ведущая роль в формировании стадии афферентного синтеза принадлежит биологическим и социальным мотивациям (потребностям, влечениям).
Следующая стадия поведения включает в себя принятие решения и формирование программ действий. Принятие решения – это, согласно Анохину, «… уменьшение числа степеней свободы», когда на основе «перебора» информации происходит формирование определенного, конкретного способа действия. Одновременно с этим формируется аппарат предвидения результатов (акцептор результатов действия).В акцепторе результатов происходит отражение свойств, предстоящих результатов в процессах опережающего возбуждения (на основании предыдущего опыта, запечатленного в элементах памяти), а также сравнение параметров реально полученных результатов с параметрами ожидаемых. При их несовпадении возникает ориентировочно-исследовательское поведение, которое приводит к формированию нового афферентного синтеза. Сличение результатов деятельности с ожидаемыми осуществляется за счет обратной афферентацииот рецепторов зрительного, слухового, тактильного, проприорецептивного и других анализаторов (Рис.2).
Биоритм – автоколебательный процесс в биологической системе, характеризующийся чередованием фаз усиления напряжения и расслабления. Выделяют микроритмы, мезоритмы, макроритмы, мегаритмы.
Стресс — это напряжение, которое возникает при появлении угрожающих или неприятных факторов в жизненной ситуации. Согласно Г. Селье выделяют три стадии стресса: стадия тревоги, стадия резистентности и стадия истощения (Рис. 1).
Дистресс – это тот стресс, который неприятен и наносит вред организму.
Стрессор
Кора большого мозга

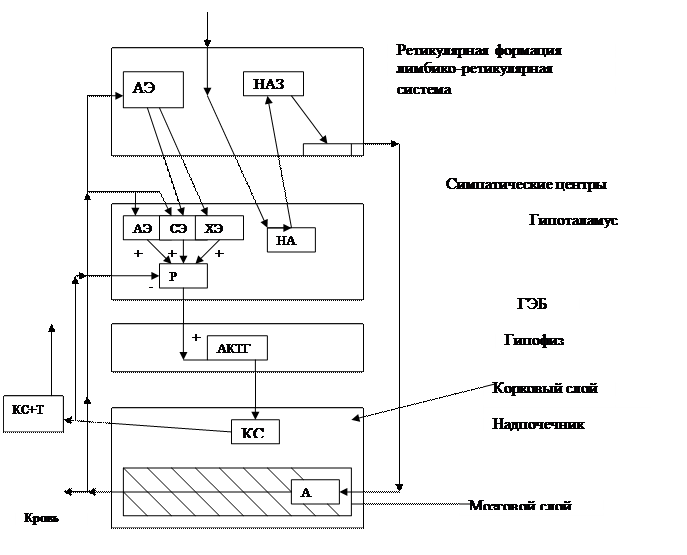
Рис. 1. Схема развития стресс-реакции (по Г.Н. Кассилю)
ГЭБ –гематоэнцефалический барьер; Р – релизинг-фактор; КС – кортикостероиды; Т – транскортин; А –адреналин; НА — норадреналин
НЕОБХОДИМО УМЕТЬ:
1. Овладеть методами исследования высших психических функций человека.
2. Научиться определять тип мыслительной деятельности человека.
3. Ознакомиться с психофизиологическим методом исследования силовых показателей в деятельности ЦНС.
4. Овладеть психологическим тестом исследования внимания и подвижности нервных процессов.
РАБОТА №7. Исследование процессов отвлечения и обобщения.
Цель работы: Познакомится с методами исследования высших психических функций человека.
Необходимо для работы: испытуемый, инструкция.
Проведение работы:
Работа проводится индивидуально каждым студентом. Необходимо к пословицам, расположенным слева, подобрать фразы или другие пословицы, сходные по смыслу, из колонки справа. Объяснить переносный смысл пословицы.
1. Куй железо, пока горячо. 1. За дровами не видно лес.
2. Когда дрова горят, тогда кашу варят.
2. Один в поле не воин. 3. Большое дело не бывает без потерь.
4. Делай все вовремя.
3. Лес рубят – щепки летят. 5. Ржавое железо не блестит.
6. Одного поля ягоды.
4. Не все золото, что блестит 7. Трудно и дереву одинокому расти.
8. Теперь обувь шьют без шила.
5. Шила в мешке не утаишь. 9. Одна пчела не много меду наносит.
10. О книге судят не по переплету.
11. Чистое золото ценится дорого.
12. Ложь не скроешь.
13. Из угольного мешка не выходит белой пыли.
14. Не по виду суди, а по делам гляди.
Понимание переносного смысла незнакомой пословицы основано на абстрагировании прямого смысла, что возможно благодаря воспроизведению временных связей, образованных в результате жизненного опыта между менее обобщающими и более обобщающими словесными обозначениями.
При непонимании переносного смысла пословиц человек подбирает выражения, имеющие лишь внешнее сходство с пословицей, что говорит о слабом абстрактном мышлении.
РАБОТА № 8. Исследование уровня развития второй сигнальной системы.
Цель работы: та же, что и в предыдущей работе.
Проведение работы:
Составить по возможности наибольшее количество слов из приведенных букв: А, Л, Й, И, А, Р, К, О, Ь, К, Л, Б.
Учитывается время выполнения задачи и количество составленных слов. Результаты исследования внести в протокол.
РАБОТА №9. Классификация понятий по «мыслительному» и «художественному» типам по (Рушкевичу)
Цель работы: научиться определять тип мыслительной деятельности человека (один из применяемых тестов).
Необходимо для работы: наборы слов-понятий.
Проведение работы:
Испытуемый должен распределить данный набор слов на три группы по три слова так, чтобы в каждой группе было что-то общее.
1. КАРАСЬ, БЕГАТЬ, ШЕРСТЬ, ОРЕЛ, ОВЦА, ПЛАВАТЬ, ЧЕШУЯ, ПЕРЬЯ, ЛЕТАТЬ.
2. ПОЕЗД, ЭЛЕКТРИЧЕСТВО, ШОФЕР, ПАР, ВАГОНОВОЖАТЫЙ, БЕНЗИН, ТРАМВАЙ, МАШИНИСТ, АВТОМОБИЛЬ.
3. СЛЫШАТЬ, ГЛАЗ, ЗАПАХ, ВИДЕТЬ, УХО, СВЕТ, НОС, ЗВУК, ОБОНЯТЬ.
4. КАРТИНА, СТАВИТЬ, КОМПОЗИТОР, ТАНЕЦ, ХУДОЖНИК, СОЧИНЯТЬ, ХОРЕОГРАФ, ПИСАТЬ, МУЗЫКА.
5. ШАХМАТЫ, ДИСТАНЦИЯ, ГРОССМЕЙСТЕР, ФУТБОЛ, СТАЙЕР, МАТЧ, БЕГ, ВРАТАРЬ, ПАРТИЯ.
6. ГНЕЗДО, МЕД, МЫШЬ, БЕРЛОГА, ВОРОБЕЙ, ХЛЕБ, ПШЕНО, МЕДВЕДЬ, НОРА.
7. РЕБЕНОК, ПРЕПОДАВАТЕЛЬ, ЯСЛИ, СТУДЕНТ, ШКОЛА, ВОСПИТАТЕЛЬ, ИНСТИТУТ, ШКОЛЬНИК, УЧИТЕЛЬ.
Работа выполняется каждым студентом индивидуально. В протокольной тетради записываются столбиком по три слова из перечисленных слов-понятий.
Анализ классификации слов производится с помощью преподавателя и на основании этого в тетрадь записывается тип мышления, преобладающий у данного студента.
РАБОТА №10. Исследование скорости ориентировочно-поисковых движений взора, а также подвижности нервных процессов.
Цель работы: познакомиться с психологическим тестом исследования внимания и подвижности нервных процессов.
Необходимо для работы: испытуемый, цифровая черно-красная таблица, секундомер.
Проведение работы:
Под вниманием понимается направленность психической деятельности, сосредоточенность ее на значимых для человека объектах. Активность внимания выражается в том, что оно носит избирательный характер. Способность человека быстро переключаться с одного вида деятельности на другой, сознательно и осмысленно перемещать внимание с одного объекта на другой называется переключением внимания.
Скорость переключения внимания у людей различна, что имеет особое значение при профессиональном отборе, так как многие профессии требуют быстрого переключения внимания.
1. Работа проводится индивидуально с одним или несколькими студентами (для сравнения).
2. Испытуемый получает задание отыскать на таблице числа, одновременно назвав их.
Работа состоит из нескольких серий заданий. В 1-й серии испытуемый называет черные числа в возрастающем порядке (от 1 до 25); во 2-й серии красные числа – в убывающем порядке (от 24 до 1); в 3-ей серии первое число черное, 24- красное, второе – черное, 23 – красное и т.д. Черные цифры отыскиваются в возрастающей, а красные – в убывающей последовательности, и так до тех пор, пока сумма пар черных и красных чисел не будет равна 25.
3. У каждого испытуемого оценивается время выполнения каждого задания (от начала поиска) и количество ошибок.
Необходимо учитывать, что в норме человек легко отыскивает как черные, так и красные цифры примерно за 45 секунд, а третье задание выполняется приблизительно за 90 секунд.
Увеличение времени работы и количества ошибок в чередовании чисел свидетельствует об инерции нервных процессов и снижении работоспособности, что может быть связано с умственным утомлением.
РАБОТА №11. Теппинг-тест
Цель работы: ознакомить студентов с психофизиологическим методом исследования силовых показателей в деятельности ЦНС.
Необходимо для работы: испытуемый, инструкция, лист белой бумаги, карандаш, секундомер.
ИНСТРУКЦИЯ по проведению теппинг-теста
Проведение работы:
1. Лист белой бумаги расчертить на шесть одинаковых прямоугольников. Испытуемый карандашом наносит с максимальной скоростью точки в первом квадрате в течение 10 с, затем во втором, в третьем и т.д. Общее время тестирования составляет 60 с.
2. Подсчитать количество точек в каждом прямоугольнике и составить шкалу по следующей схеме.
| Время (с) | ||||||
| Количество точек |
3. На основании результатов сделать выводы о силе и устойчивости процесса
возбуждения. Оценку производить совместно с преподавателем.
РАБОТА №12. Исследование функциональной ассиметрии мозга
Цель работы: с помощью функциональных проб определить латеральный фенотип человека.
Необходимо для работы: секундомер, динамометр, лист бумаги с отверстием диаметром 1 см.
Проведение работы:
1. Студенты по двое проводят следующие пробы для оценки сенсомоторных ассиметрий.
| Пробы | Оцениваемые показатели |
| 1. Взятие ручки со стола | Ведущей считается рука, которой испытуемый берет предмет |
| 2. Без зрительного контроля одновременно двумя руками рисуются круги, квадраты, треугольники | Ведущей считается рука, которой более эффективно осуществляется движение |
| 3. Переплетение пальцев рук. | Большой палец ведущей руки ложится сверху |
| 4. Поза Наполеона (скрести руки на груди) | Ведущая рука первой начинает движение и располагает кисть на противоположном предплечье. |
| 5. Динамометрия | Трижды определяется сила сжатия динамометра каждой рукой. Определяются средние значения. Большая сила выявляется у ведущей руки. |
| 6. Аплодирование. | Ведущей считается более активная в движении рука. |
| 7. Тест вытянутых рук. С закрытыми глазами обе руки вытягиваются вперед | Ведущей считается поднятая выше рука. |
| 8. Закидывание ноги на ногу. | Ведущая нога располагается сверху. |
| 9. Подпрыгивание на одной ноге | Ведущей считается толчковая нога. |
| 10. Приближение к двери, возвращение пятясь | Ведущая нога начинает движение. |
| 11. Проба с секундомером. | Ведущим считается ухо, которое испытуемый приближает к секундомеру. |
| 12. Проба с секундомером. | Ведущим считается ухо, которое громче слышит тиканье часов. |
| 13. Проба “Дырка в карте”. Испытуемый фиксирует предмет через небольшое отверстие в листе бумаги. Затем поочередно закрывает правый и левый глаза | Закрытие ведущего глаза ведет к исчезновению предмета из поля зрения. |
| 14. Поочередное прищуривание глаза. | 14. Первым прищуривается ведущий глаз глаза. |
2. Результаты проб студенты заносят в протокол.
Протокол эксперимента по определению латерального фенотипа.
| П | ||||||||||||||
| Л |
3. На основании функциональных проб определяют по формуле коэффициент латерализации:
(П-Л)/(П+Л) • 100%, где П – количество правосторонних признаков, Л — количество левосторонних признаков.
4. На основании коэффициента латерализации определяют моторный фенотип (пробы 1-10), сенсорный (пробы 11-15) и общий сенсомоторный фенотип (по всем пробам).
5. По коэффициенту латерализации (положительный – правосторонняя, отрицательный — левосторонняя) определяют ведущее полушарие в моторной и сенсорной деятельности.