Научение и память.
Одним из биологических аспектов психики является выработка новых форм реагирования на воздействия, семантическая значимость которых меняется или с которыми человек вообще раньше не сталкивался. Эту способность принято обозначать как научение, которое можно определить как совокупность процессов, обеспечивающих выработку и закрепление форм реагирования, адекватных физиологическим, биологическим и социальным потребностям. Следует иметь в виду, что это термин комплексный, объединяющий два понятия: обучение, где присутствуют обучающий и формы обучения, а также учение – обучаемый и условия обучения. С точки зрения психофизиологии, т. е. процессов и механизмов, обеспечивающих научение, это также явление комплексное, включающее потребность к научению, т. е. мотивационно эмоциональную сферу; внимание как непроизвольное, так и произвольное; восприятие; память; мышление; соотношение сознательного и бессознательного; автоматизацию навыков и некоторые другие.
Принято различать три группы способов (механизмов) научения по степени участия в них организма как целого: 1) реактивное поведение; 2) оперантное поведение (или научение в результате оперантного обусловливания) и 3) когнитивное научение.
Реактивное поведение проявляется в том, что организм реагирует пассивно, но при этом трансформируются нейронные цепи и формируются новые следы памяти. Среди разновидностей реактивного поведения различают: а) привыкание; б) сенсибилизацию; в) импринтинг и г) условные рефлексы. Привыкание (или габитуация) заключается в том, что организм в результате изменений на уровне рецепторов или ретикулярной формации «научается» игнорировать какой то повторный или постоянно действующий раздражитель, «убедившись», что он не имеет особого значения для той деятельности, которая в данный момент осуществляется. Сенсибилизация представляет собой процесс противоположный. Повторение стимула ведет к более сильной активации организма, который становится все более и более чувствительным к данному стимулу. Импринтинг (запечатление) – наследственно запрограммированное и необратимое формирование определенной специфической формы реагирования, например привязанность новорожденных животных к первому движущемуся объекту, который попадет в его поле зрения в первые часы жизни. Условные рефлексы, или классическое обусловливание, ассоциативное обусловливание, по И. П. Павлову, – основной механизм индивидуального приспособления организма.
Артифициальные стабильные функциональные связи (АСФС) представляют собой закрепление в долговременной памяти связи между фармакологическим и физическим (фотостимуляция) эффектами после одноразового их сочетания.
Презентация на тему: Психика и организм
... следить за несколькими процессами одновременно),переключение. а) природное, связанное с саморегуляцией организма (непроизвольное); б) социально обусловленное, связанное с воспитанием и обучением ( ... альтруисты, любят помогать другим, нерешительны). Воля – сознательное регулирование человеком своего поведения, связанное с преодолением внутренних и внешних препятствий. Эмоции – своеобразное выражение ...
Оперантное поведение, или научение в результате оперантного обусловливания, представляет собой закрепление тех действий, последствия которых для организма желательны, и отказ от действий, приводящих к нежелательным последствиям. Различают три разновидности этого типа научения: а) метод проб и ошибок; б) формирование автоматизированных реакций и в) подражание. Научение методом проб и ошибок заключается в том, что, перебирая способы достижения цели (преодоления препятствий), человек отказывается от неэффективных и в конце концов находит решение задачи. Формирование автоматизированных реакций – это создание очень сложных поведенческих реакций поэтапно. Каждый этап при этом подкрепляется (положительное и отрицательное подкрепление, угасание, дифференцировка, генерализация).
На этой основе была выдвинута концепция программированного обучения, первоначальное увлечение которой сменилось разочарованием ввиду низкой эффективности этого метода обучения. Подражание представляет собой научение путем наблюдения и воспроизведения действий модели, причем их значение не всегда понимается. Оно свойственно в основном приматам. Различают две формы подражания: чистое подражание и викарное научение, т. е. научение с пониманием.
Когнитивное научение в эволюционном отношении наиболее поздний и наиболее эффективный тип научения. В полном объеме такое научение присуще только людям, хотя какие то его эволюционные предшественники или отдельные элементы мы можем выделить и у высших животных. Различают следующие формы когнитивного научения: а) латентное научение; б) обучение сложным психомоторным навыкам; в) инсайт и г) научение путем рассуждений. Латентное научение – аналитическая обработка поступающей информации, а также уже имеющейся (хранящейся) в памяти и на этой основе выбор адекватной реакции. Обучение сложным психомоторным навыкам, которыми человек на протяжении своей жизни овладевает в большом объеме в зависимости от индивидуальных особенностей организации психомоторной активности, образа жизни, профессии и т. п., проходит через стадию когнитивной стратегии (выбор программы), ассоциативную (проверка и совершенствование этой программы) и автономную стадии, когда психомоторный навык переходит на уровень автоматизма с ослаблением или полным отсутствием контроля сознания. Инсайт (от англ. insight – прозрение, проникновение; во французском языке идентичный ему термин – интуиция) заключается в том, что информация, «разбросанная» в памяти, как бы объединяется и используется в новой интеграции. Человеку кажется, что решение приходит спонтанно, хотя на самом деле, конечно, это не так, а скорее всего результат подсознательной аналитико синтетической деятельности. Научение путем рассуждений – это научение посредством мыслительного процесса. Фундаментом для мышления служит перцептивное научение (опознание образа) и концептуальное научение (абстрагирование и обобщение).
Для отдельных форм научения в процессе развития существуют критические периоды, когда организм наиболее чувствителен к этим формам. Один из наиболее ярких примеров – первичное усвоение языка. Некоторые виды поведения, информация, усвоенные в каком то особом состоянии сознания, могут не проявляться в состоянии активного бодрствования, но проявляются вновь, когда организм возвращается в это специфическое состояние (например, сомнамбулизм, гипноз, под воздействием некоторых психотропных веществ).
Научение и память
... бы быть структурной основой некоторых видов научения и памяти. Но окончательные выводы сделать пока невозможно. Гиппокамп получает информацию от всех органов чувств. Сигналы, идущие ... Пластичность головного мозга: воздействие окружающей среды. .13 10. Литература. …………………………………………………………..15 НАУЧЕНИЕ И ПАМЯТЬ. Когда экспериментаторы предотвращают воздействие внешних стимулов на ту или иную сенсорную ...
В процессе обучения могут развиваться различные формы взаимодействия с ранее усвоенными знаниями и навыками, в частности явление переноса – облегчение обучения на основе ранее приобретенных опыта и знаний и противоположное ему затруднение при перестройке, переделке ранее очень прочно усвоенных стереотипов.
Механизмы научения весьма разнообразны по характеру физиологических процессов и вовлекаемых структур нервной системы. На уровне нейрона это проявляется в изменении уровня поляризации мембран – длительная деполяризация или гиперполяризация. На уровне межнейронального взаимодействия – в изменении активности кальциевых каналов, что приводит к изменению медиаторной активности, росте синаптических терминалий, изменении состояния синаптических структур и происходящих в них процессов, особенно касающихся ацетилхолина и глутамата. Среди структур мозга, имеющих непосредственное отношение к процессам научения (скорость, объем, эффективность), в первую очередь следует выделить неспецифическую активирующую систему мозга, образования лимбической системы (гиппокамп, миндалины), лобно височные отделы мозга и другие ассоциативные зоны коры с учетом функциональной специализации правого и левого полушарий. У правшей усвоение абстрактно логической информации связано в большей степени с левым полушарием, а наглядно образная, эмоциональная окраска – с правым. Среди факторов, влияющих на научение человека, существенное значение имеют возраст,мотивация, состояние таких психических функций, как внимание, память, мышление и др., а также индивидуальные особенности (способности).
Таким образом, проблема научения является одной из фундаментальных в психологии вообще и психофизиологии в частности, поскольку она позволяет понять механизм психической адаптации человека к условиям существования, сколь бы они ни были необычными, своеобразными, чрезвычайными.
В непосредственной связи с научением находится проблема памяти. В поведенческой активности они, безусловно, составляют единое целое. Но вместе с тем отождествлять их нельзя, а в дидактическом отношении память тем более требует самостоятельного рассмотрения.
Память представляет собой совокупность процессов, обеспечивающих восприятие, запечатление, хранение и воспроизведение (извлечение) информации. Поскольку последний компонент нередко выступает как критерий памяти, то к рассматриваемой совокупности следует отнести и забывание, под которым понимается либо безвозвратная утрата информации, либо невозможность ее извлечения при обычных условиях. Существует много подходов к классификации видов памяти. Для нас наиболее существенное значение представляет временная характеристика сохранения способности к воспроизведению запечатленной информации. С этой точки зрения различают следующие виды памяти.
Сенсорная (иконическая, следовая) память, которая обеспечивает сохранение воспринятого образа на протяжении долей секунды. Кратковременная (первичная) память позволяет удерживать воспринятую информацию на протяжении около 20 с. Долговременная (вторичная) память простирается на очень большой временной диапазон, начиная от 20 с и простираясь на минуты, часы, дни, месяцы, годы. По всей видимости, это понятие включает несколько существенно различных видов памяти. Так, в частности, во вторичной памяти выделяют оперативную память, т. е., по существу, пролонгированную кратковременную память. Суть ее заключается в том, что под влиянием внутренней мотивации или каких либо внешних обстоятельств способность к воспроизведению продлевается на промежуток времени более 20 с.
Образ вожатого
Хороший вожатый — тот, у которого в отряде всегда хорошие дети (такое впечатление, что ему просто везет), мягкий, дружеский климат в коллективе. Его радует каждая предстоящая встреча с ребятами. Хороший вожатый всегда готов: играть с детьми, гулять, петь, выступать со сцены, сочинять, рисовать, шутить.., а главное — чутко реагировать на настроение ребят, на ситуацию в отряде, как шахматист ...
Выделяют еще так называемую «вечную», или третичную, память, когда способность воспроизводить когда то запечатленную информацию сохраняется на протяжении всей оставшейся жизни (например, имена свое и ближайших родственников и т. п.).
И только в патопсихологических ситуациях разрушается и этот вид памяти.
Каждую из перечисленных форм памяти характеризуют также по объему запечатлеваемой информации. По этому показателю колебания оказываются очень существенными. Так, для сенсорной памяти этот объем весьма велик и ограничивается, по существу, информационной емкостью самого воспринимаемого объекта. Однако лишь только небольшая часть переходит в память кратковременную. Для большинства людей объем кратковременной памяти составляет 7 ± 2 блока информации, а вот объем блока может существенно колебаться в зависимости от индивидуальных особенностей человека и главным образом от уже хранящейся у него информации в долговременной памяти. Чем больше человек знает, тем крупнее у него эти блоки. Таким образом, оказывается весьма различным объем кратковременной памяти, измеряемой в тех или иных элементарных единицах. Для долговременной памяти характерен практически не ограниченный ее потенциальный объем, заполнение которого до предела просто невозможно на протяжении реальной жизни человека. Таким образом, возможности запечатления новой информации у человека безграничны.
Различные виды памяти имеют весьма отличные физиологические механизмы. Так, в частности, сенсорная (следовая) память может быть, по существу, идентифицирована с последовательными образами, развитие которых характерно практически для всех сенсорных систем. Наиболее отчетливо они проявляются в зрительном анализаторе, поэтому они хорошо известны практически всем людям. Длительность такого образа зависит от интенсивности раздражителя и степени контраста и сохраняется порой на несколько секунд. Существование последовательных образов обусловлено постепенно затухающими следовыми процессами в изменении уровня поляризации мембран рецепторно нейрональных структур анализатора. Если условия складываются таким образом, что информация не переходит в регистры кратковременной или долговременной памяти, то эти следовые процессы безвозвратно исчезают, особенно когда воздействует новый сигнал. Основой долговременной памяти является формирование энграмм – структурно функциональных комплексов запечатления информации. Для образования таких энграмм непременным условием является достаточно длительная реверберация сигналов, связанных с информацией, находящейся в регистре первичной (кратковременной) памяти. Пролонгации реверберации способствуют следующие факторы: во первых, это неоднократное повторение воспринимаемой информации; во вторых, осмысливание этой информации, установление ее логической структуры или связи с уже хранящейся в долговременной памяти информацией, что резко сокращает количество необходимых повторений; в третьих, установка на длительное запоминание; в четвертых, высокий интерес к запоминаемому материалу, что даже без повторного восприятия значительно увеличивает длительность реверберации.
Память и её виды 2
... прекращения этого воздействия. Они сохраняются и вотсутствии этих предметов и явлений в виде представлений памяти. Важнейшая особенностьпсихики состоит в том, что отражение внешних воздействий постоянно используетсяиндивидом ... в кладовых его мозга. Наш мозг обладает важнымсвойством. Он не только получает информацию об окружающем мире, но и хранитнакапливает её. Образцы предметов иявлений, которые ...
Кроме того, существенное значение имеет функциональное состояние организма, степень утомления. Всем известно, что «на свежую голову» запоминать легче, однако для образования энграмм необходим сон, особенно быстрый сон. На процессы закрепления информации в долговременной памяти существенное влияние оказывает эмоциональный фон, связанный с этой информацией. Гораздо лучше запоминаются факты, имеющие положительную эмоциональную окраску, хуже – отрицательную и еще хуже – не имеющие никакой эмоциональной окраски. Имеются некоторые достижения в улучшении памяти фармакологическими средствами, которые, однако, используются только в медицинской практике, но не в повседневной жизни.
Механизм самих энграмм понят не до конца. В настоящее время можно говорить о перестройках на уровне синапса, включая все звенья химической передачи, т. е. и синтез медиатора, и участие модуляторов, и свойства постсинаптической мембраны, особенно расположенных на ней фармакологических рецепторов. Представляется безусловной значимость нуклеиновых кислот (ДНК – дезоксирибо нуклеиновой кислоты, РНК – рибонуклеиновой кислоты), которые обладают неисчерпаемыми возможностями химического кодирования как генетически закрепленной, так и приобретаемой в процессе индивидуальной жизнедеятельности информации. Не исключена возможность и некоторых структурных изменений, в частности разрастание пресинаптических терминалей или активизация «латентных» (недействующих) синапсов. Нет полной определенности относительно конкретного местонахождения энграмм. В настоящее время наиболее распространенной является точка зрения, что «хранилища» долговременной памяти представляют собой в морфофункциональном отношении свойство всей ассоциативной коры.
Медицинская практика показывает, что нет таких ограниченных участков высших отделов мозга, поражение которых приводило бы к катастрофическим расстройствам памяти, в то время как диффузные поражения значительной массы мозга по достижении определенной степени (как это нередко встречается у весьма пожилых людей) приводят к ослаблению сначала кратковременной, затем оперативной, долговременной и даже вечной памяти.
Процесс забывания следует рассматривать в двух аспектах. Во первых, забывание может быть по своей природе интерпретировано как «стирание» следов ранее хранимой информации. Для первичной памяти – это главным образом интерференция, получение новой информации. Для вторичной и третичной памяти, которые характеризуются наличием энграмм, такого рода забывание обусловлено их разрушением в результате тех или иных воздействий. Во вторых, забывание может быть связано с затруднением, а иногда и полной неспособностью к воспроизведению при сохранных энграммах. И только специальными способами (например, гипноз, некоторые фармакологические средства, электрическое раздражение при нейрохирургических операциях и т. п.) можно убедиться, что полностью информация не утрачена.
- 7.1. Классификация видов памяти
- 7.2. Физиологические теории памяти
- 7.3. Биохимические исследования памяти
Одним из основных свойств нервной системы является способность к длительному хранению информации о внешних событиях. По определению, память — это особая форма психического отражения действительности, заключающаяся в закреплении, сохранении и последующем воспроизведении информации в живой системе. По современным представлениям, в памяти закрепляются не отдельные информационные элементы, а целостные системы знаний, позволяющие всему живому приобретать, хранить и использовать обширный запас сведений в целях эффективного приспособления к окружающему миру. Память как результат обучения связана с такими изменениями в нервной системе, которые сохраняются в течение некоторого времени и существенным образом влияют на дальнейшее поведение живого организма. Комплекс таких структурно-функциональных изменений связан с процессом образования энграмм — т.е. следов памяти (термин, предложенный зоологом Дж. Янгом в 50-х гг.).
Психофизиология памяти 3
... периода, – это этап долговременной памяти. Фиксация энграммы осуществляется при помощи процесса консолидации. Консолидация начинает развиваться во время пребывания следа в фазе кратковременного ... заранее оговоренного срока. Срок хранения информации колеблется от нескольких секунд до нескольких дней; долговременная память— память, способная хранить информацию в течение неограниченного срока. ...
Память выступает также как своеобразный информационный фильтр, поскольку в ней обрабатывается и сохраняется лишь ничтожная доля от общего числа раздражителей, воздействующих на организм. Без отбора и вытеснения информации из памяти живое существо было бы, образно говоря, «затоплено» бесконечным потоком поступающих извне раздражителей. Результаты этого были бы так же катастрофичны, как и отсутствие способности к обучению и памяти.
7.1. Классификация видов памяти
- 7.1.1. Элементарные виды памяти и научения
- 7.1.2. Специфические виды памяти
- 7.1.3. Временная организация памяти
- 7.1.4. Механизмы запечатления
Пронизывая все стороны существования человека, память имеет разные формы и уровни проявления и функционирования.
7.1.1. Элементарные виды памяти и научения
В нейрофизиологии выделяют следующие элементарные механизмы научения: привыкание, сенситизацию, временную связь (условный рефлекс).
Согласно И.П. Павлову, физиологической основой запоминания служит условный рефлекс как акт образования временной связи между стимулом и реакцией. Эти формы памяти и научения называют простыми, чтобы отличать от научения, имеющего произвольный, осознанный характер. Элементарные формы научения есть даже у беспозвоночных. Привыкание проявляется в постепенном уменьшении реакции по мере повторного предъявления раздражителя. Привыкание всегда сопровождает угашение ориентировочной реакции. Сенситизация — это процесс, противоположный привыканию. Он выражается в снижении порога при предъявлении раздражителей. Благодаря сенситизации организм начинает реагировать на ранее нейтральный раздражитель. Существует также разделение памяти на генотипическую и фенотипическую. Первая — генотипическая, или филогенетическая, связана с безусловными рефлексами и инстинктами. Вторая — фенотипическая — обеспечивает обработку и хранение информации, приобретаемой в ходе онтогенеза на основе различных механизмов научения. 
7.1.2. Специфические виды памяти
В ходе совершенствования механизмов адаптации развились и упрочились более сложные формы памяти, связанные с запечатлением разных сторон индивидуального опыта.
Психофизиология памяти
... цепях нейронов, а долговременная память связана с морфофункциональными изменениями устойчивого характера, происходящими в синапсах, увеличивающими или уменьшающими их проводимость. Из кратковременной в долговременную память информация переходит ... а также с содержанием РНК в тех или иных образованиях мозга. В коре головного мозга следы памяти или научения обнаруживаются в виде изменений в клетках ц. ...
Модально-специфические виды. Мнестические процессы могут быть связаны с деятельностью разных анализаторов, поэтому существуют специфические виды памяти соответственно органам чувств: зрительная, слуховая, тактильная, обонятельная, двигательная. Следует упомянуть, что уровень развития этих видов памяти у разных людей различен. Не исключено, что последнее связано с индивидуальными особенностями анализаторных систем. Например, встречаются индивиды с необыкновенно развитой зрительной памятью. Это явление — эйдетизм — выражается в том, что человек в нужный момент способен воспроизвести во всех деталях ранее виденный предмет, картину, страницу книги и т.д. Эйдетический образ отличается от обычных тем, что человек как бы продолжает воспринимать образ в его отсутствие. Предполагается, что физиологическую основу эйдетических образов составляет остаточное возбуждение зрительного анализатора. Хорошо развития модально-специфическая память нередко является профессионально важным качеством: например, слуховая память музыкантов, вкусовая и обонятельная дегустаторов, двигательная гимнастов и т.д.
Образная память. Запечатление и воспроизведение картин окружающего мира связаны с синтезом модально-специфических впечатлений. В этом случае фиксируются сложные образы, объединяющие зрительные, слуховые и другие модально-специфические сигналы. Такую память называют образной. Образная память гибка, спонтанна и обеспечивает длительное хранение следа. По некоторым представлениям, ее морфологической основой служат сложные нейрональные сети, включающие взаимосвязанные нейронные звенья, расположенные в разных отделах мозга. Поэтому выпадение какого-либо одного звена или нескольких звеньев образной памяти не способно разрушить всю ее структуру. Это дает образной памяти большие преимущества как в эффективности процессов усвоения и хранения, так и в объеме и прочности фиксации информации. Вероятно, что с подобными особенностями образной памяти связаны внезапные, нередко безо всяких усилий припоминания забытого материала. Помимо этого иногда выделяют также эмоциональную и словесно-логическую память.
Эмоциональная память. Эмоциональная память связана с запоминанием и воспроизведением эмоциональных переживаний. Эмоционально окрашенные воспоминания могут возникать как при повторном воздействии раздражителей, обусловивших это состояние, так и в отсутствие последних. Эмоционально окрашенное впечатление фиксируется практически мгновенно и непроизвольно, обеспечивая пополнение подсознательной сферы человеческой психики. Так же непроизвольно информация воспроизводится из эмоциональной памяти. Этот вид памяти во многом сходен с образной, но иногда эмоциональная память оказывается даже более устойчивой, чем образная. Ее морфологической основой предположительно служат распределенные нервные сети, включающие нейрональные группы их разных отделов коры и ближайшей подкорки.
Функциональная классификация нейронов
... находиться долгое время в активности. Для этого в префрональную кору информация переписывается – там найдены такие же нейроны. Найдены участки, активирующиеся на неживые объекты (здания, инструменты и ... различных зон. Регистрация нейронной и мультинейронной активности –вживляют или вводят микроэлектроды в мозг – помогает находить очаги возбуждения при эпилепсии и болезни Паркинсона. Регистрация 1 ...
Словесно-логическая память. Словесно-логическая (или семантическая) — это память на словесные сигналы и символы, обозначающие как внешние объекты, так и внутренние действия и переживания. Ее морфологическую основу можно схематически представить как упорядоченную последовательность линейных звеньев, каждое из которых соединено, как правило, с предшествующим и последующим. Сами же цепи соединяются между собой только в отдельных звеньях. В результате выпадение даже одного звена (например, вследствие органического поражения нервной ткани) ведет к разрыву всей цепи, нарушению последовательности хранимых событий и к выпадению из памяти большего или меньшего объема информации.
7.1.3. Временная организация памяти
- Другим основанием для классификации памяти является продолжительность закрепления и сохранения материала. Принято подразделять память на три вида:
- иконическую, или сенсорную, память (ИП);
- кратковременную, или оперативную, память (КВП);
- долговременную, или декларативную, память (ДВП).
Иногда последний вид памяти называется пропозиционной, вторичной или семантической. Считается, что каждый из этих видов памяти обеспечивается различными мозговыми процессами и механизмами, связанными с деятельностью функционально и структурно различных мозговых систем. Длительность хранения в сенсорной, или иконической, памяти составляет 250-400 мс, однако по некоторым данным этот процесс может продолжаться до 4-х сек. Объем ИП при наличии соответствующей инструкции от 12 до 20 элементов. Длительность хранения в кратковременной памяти около 12 сек., при повторении дольше. Объем КВП представлен широко известным числом Миллера 7 ± 2 элемента. Длительность хранения в ДВП неопределенно долгая, объем велик, по некоторым представлениям, неограничен. Подобную временную типологию памяти подтверждают эксперименты с животными по научению, в которых показано, что запоминание ухудшается, если сразу же за научением следует удар электрическим током (электроконвульсивный шок — ЭКШ), т.е. ЭКШ препятствует переносу информации из кратковременной памяти в долговременную. Аналогично, травма, полученная человеком, не сказывается сразу на воспроизведении событий, но уже через несколько минут человек не может точно вспомнить всех обстоятельств происшествия. О существовании двух различных хранилищ памяти (долговременного и кратковременного) свидетельствуют такие факты. Две группы испытуемых — здоровые и больные амнезией — должны были воспроизвести список из 10 слов сразу после заучивания и с задержкой в 30 сек. В момент задержки испытуемые обеих групп должны были решать арифметическую задачу. Значимых различий между двумя группами испытуемых при немедленном воспроизведении обнаружено не было, в то время как при отсроченном воспроизведении у больных амнезией объем запоминания был намного ниже. Этот эксперимент подтверждает, что механизмы кратковременной и долговременной памяти у человека различны.
- Во всех вышеперечисленных видах памяти имеет место фиксация информации, включающая в себя, по крайней мере, три этапа:
- формирование энграммы, т.е. следа, оставляемого в мозгу тем или иным событием;
- сортировка и выделение новой информации;
- долговременное хранение значимой информации.
7.1.4. Механизмы запечатления
Сложной проблемой является механизм образования следов памяти, выделение структурных образований, участвующих в хранении и воспроизведении имеющихся следов, а также тех структур, которые регулируют эти процессы.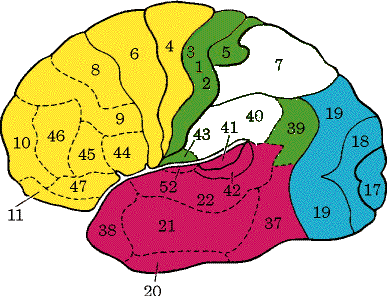
Память как психический процесс 4
... путей принятия, переработки, хранения информации машинами. Это потребовало технического и алгоритмического моделирования процессов памяти по аналогии с человеческим мозгом. Психологические теории памяти. В этой группе теорий ... и, соответственно, свои теории. Физическая теория памяти. Согласно этой теории нервный импульс, проходя через определенную группу нейронов (нервных клеток), вызывает в местах ...
Опыты К. Лешли. Пионер в области исследования памяти Карл Лешли пытался с помощью хирургического вмешательства в мозг дать ответ о пространственном расположении памяти, по аналогии с речевыми, моторными или сенсорными зонами. Лешли обучал разных животных решать определенную задачу. Потом он удалял у этого животного один за другим различные участки коры — в поисках места расположения следов памяти — энграмм. Однако, независимо от того, какое количество корковой ткани было удалено, найти то специфическое место, где хранятся следы памяти (энграммы) Лешли не удалось. Свою классическую статью он закончил выводом о том, что память одновременно находится в мозгу везде и нигде. Впоследствии этим фактам было найдено объяснение. Оказалось, что в процессах памяти участвуют не только кора, но многие подкорковые образования и, кроме того, следы памяти широко представлены в коре и при этом многократно дублируются.
Этапы формирования энграмм. По современным представлениям, фиксация следа в памяти осуществляется в три этапа. Вначале, в иконической памяти на основе деятельности анализаторов возникают сенсорный след (зрительный, слуховой, тактильный и т. п.).
Эти следы составляют содержание сенсорной памяти. На втором этапе сенсорная информация направляется в высшие отделы головного мозга. В корковых зонах, а также в гиппокампе и лимбической системе происходит анализ, сортировка и переработка сигналов, с целью выделения из них новой для организма информации. Есть данные, что гиппокамп в совокупности смедиальной частью височной доли играет особую роль в процессе закрепления (консолидации) следов памяти. Речь идет о тех изменениях, которые происходят в нервной ткани при образовании энграмм. Гиппокамп, по-видимому, выполняет роль селективного входного фильтра. Он классифицирует все сигналы и отбрасывает случайные, способствуя оптимальной организации сенсорных следов в долговременной памяти. Он также участвует в извлечении следов из долговременной памяти под влиянием мотивационного возбуждения. Роль височной области предположительно состоит в том, что она устанавливает связь с местами хранения следов памяти в других отделах мозга, в первую очередь, в коре больших полушарий. Другими словами, она отвечает за реорганизацию нервных сетей в процессе усвоения новых знаний; когда реорганизация закончена, височная область в дальнейшем процессе хранения участия не принимает. На третьем этапе следовые процессы переходят в устойчивые структуры долговременной памяти. Перевод информации из кратковременной памяти в долговременную по некоторым предположениям может происходить как во время бодрствования, так и во сне.
Системы регуляции памяти. Важным параметром классификации памяти является уровень управления, или регуляции, мнестических процессов. По этому признаку выделяют непроизвольную и произвольную память. В первом случае запоминание и воспроизведение происходит без усилий, во втором — в результате осознанной мнестической деятельности. Очевидно, что эти процессы имеют разное мозговое обеспечение. В целом система управления и регуляции памяти в головном мозге включает неспецифические и специфические компоненты. При этом выделяются два уровня регуляции: 1) неспецифический (общемозговой) — сюда относят ретикулярную формацию, гипоталамус, неспецифический таламус, гиппокамп и лобную кору; 2) модально-специфический (локальный), связанный с деятельностью анализаторных систем. По современным представлениям, неспецифический уровень регуляции участвует в обеспечении практически всех видов памяти. Из клиники очаговых поражений мозга известно, что существуют так называемые модально-неспецифические расстройства памяти, когда ослабление или утрата функций памяти не зависит от характера стимула. Они возникают при поражении глубоких структур мозга: ретикулярной формации ствола, диэнцефальной области, лимбической системы, гиппокампа. В случае поражения гиппокампа возникает известное заболевание — корсаковский синдром, при котором больной при сравнительной сохранности следов долговременной памяти утрачивает память на текущие события. Установлено также, что при активации ретикулярной формации формирование энграмм происходит эффективнее, а при снижении уровня активации, напротив, ухудшается как непроизвольное, так и произвольное запоминание любого нового материала, независимо от его сложности и эмоциональной значимости. Наряду с этим улучшение кратковременной памяти (увеличение объема при предъявлении информации в быстром темпе) может наблюдаться при электрической стимуляции таламокортикальной системы. В то же время при разрушении ряда областей таламуса возникают затруднения в усвоении новой информации или сохранении заученной ранее. В обеспечении произвольного запоминания, или мнестической деятельности, ведущую роль играют лобные доли коры, особенно левой лобной доли. Модально-специфический, или локальный уровень, регуляции памяти обеспечивается деятельностью анализаторных систем, главным образом на уровне первичных и ассоциативных зон коры. При их нарушении возникают специфические формы нарушения мнестических процессов, имеющие избирательный характер. Из сказанного следует, что система регуляции памяти имеет иерархическое строение, и полное обеспечение функций и процессов памяти возможно лишь при условии функционировании всех ее звеньев. Память следует понимать как системное (эмерджентное) свойство всего мозга и даже целого организма. Поэтому уровень, на котором возможно понимание памяти, — это уровень живой системы в целом (см. Хрестомат. 7.2).
7.2. Физиологические теории памяти
В современной нейробиологии и психофизиологии существует целый ряд теорий и моделей, объясняющих разные стороны функционирования памяти.
Теория Д. Хебба. Первые исследования физиологических основ памяти связаны с именем Д. Хебба. В 40-е гг. он ввел понятия кратковременной и долговременной памяти и предложил теорию, объясняющую их нейрофизиологическую природу. По Хеббу, кратковременная память — это процесс, обусловленный повторным возбуждением импульсной активности в замкнутых цепях нейронов, не сопровождающийся морфологическими изменениями. Долговременная память, напротив, базируется на структурных изменениях, возникающих в результате модификации межклеточных контактов — синапсов. Хебб полагал, что эти структурные изменения связаны с повторной активацией (по его определению — «повторяющейся реверберацией возбуждения») замкнутых нейронных цепей, например, путей от коры к таламусу или гиппокампу и обратно к коре. Повторное возбуждение нейронов, образующих такую цепь, приводит к тому, что в них возникают долговременные изменения, связанные с ростом синаптических соединений и увеличением площади их контакта между пресинаптическим аксоном и постсинаптической клеточной мембраной. После установления таких связей эти нейроны образуют клеточный ансамбль, и любое возбуждение хотя бы одного относящегося к нему нейрона, приводит в возбуждение весь ансамбль. Это и есть нейрональный механизм хранения и извлечения информации из памяти. Непосредственно же основные структурные изменения, согласно Хеббу, происходят в синапсах в результате процессов их роста или метаболических изменений, усиливающих воздействие каждого нейрона на следующий нейрон. Достоинство этой теории в том, что она толкует память не как статическую запись или продукт изменений в одной или нескольких нервных клетках, а как процесс взаимодействия многих нейронов на основе соответствующих структурных изменений. Современные подходы к изучению физиологических механизмов памяти в значительной степени связаны с развитием изложенных выше идей Д. Хебба.
Синаптическая теория. Свое название эта теория получила из-за того, что главное внимание в ней уделяется роли синапса в фиксации следа памяти. Она утверждает, что при прохождении импульса через определенную группу нейронов возникают стойкие изменения синаптической проводимости в пределах определенного нейронного ансамбля. Один из наиболее авторитетных исследователей нейробиологических основ памяти, С. Роуз подчеркивает: при усвоении нового опыта, необходимого для достижения каких-либо целей, происходят изменения в определенных клетках нервной системы. Эти изменения, выявляемые морфологическими методами с помощью световой или электронной микроскопии, представляют собой стойкие модификации структуры нейронов и их синаптических связей. Г. Линч и М. Бодри (1984) предложили следующую гипотезу. Повторная импульсация в нейроне, связанная с процессом запоминания, предположительно, сопровождается увеличением концентрации кальция в постсинаптической мембране, что приводит к расщеплению одного из ее белков. В результате этого освобождаются замаскированные и ранее неактивные белковые рецепторы (глутаматрецепторы).
За счет увеличения числа этих рецепторов возникает состояние повышенной проводимости синапса, которое может сохраняться до 5-6 суток. Эти процессы тесно связаны с увеличением диаметра и усилением активности так называемого аксошипикового синапса — наиболее пластичного контакта между нейронами. Одновременно с этим образуются новые шипики на дендритах, а также увеличиваются число и величина синапсов. Таким образом, экспериментально показаны морфологические изменения, сопровождающие формирование следа памяти.
Реверберационная теория. Основания теории были выдвинуты известным нейрофизиологом Л. де Но. Теория базировалась на существовании в структурах мозга замкнутых нейронных цепей. Известно, что аксоны нервных клеток соприкасаются не только с дендритами других клеток, но могут и возвращаться обратно к телу своей же клетки. Благодаря такой структуре нервных контактов, появляется возможность циркуляции нервного импульса по реверберирующим (постепенно затухающим) кругам возбуждения разной сложности. В результате возникающий в клетке разряд возвращается к ней либо сразу, либо через промежуточную цепь нейронов и поддерживает в ней возбуждение. Эти стойкие круги реверберирующего возбуждения не выходят за пределы определенной совокупности нервных клеток и рассматриваются как физиологический субстрат сохранения энграмм. Именно в реверберационном круге возбуждения происходит переход из кратковременной в долговременную память. С этим непосредственно связана гипотеза А.С. Батуева о двух нейронных системах, обеспечивающих оперативную память. Одна система, включающая «нейроны памяти», работает на эстафетно-реверберационном принципе передачи информации, когда отдельные группы нейронов памяти вовлекаются друг за другом, представляя собой своеобразные «нейронные ловушки», поскольку возбуждение в них циркулирует в течение 1,5-2 с. Другая система обеспечивает надежность переходных процессов: переключение информации с «сенсорных» нейронов на «нейроны памяти» и далее на нейроны «моторных программ» и т.д. Их взаимодействие позволяет эффективно запоминать текущую информацию. Однако реверберационная теория не дает ответа на ряд вопросов. В частности, она не объясняет причину возврата памяти после электрошоковых воздействий, когда, согласно этой теории, в подобных случаях возврата памяти не должно быть.
Нейронные модели памяти. С развитием микроэлектродной техники появилась возможность изучения электрофизиологических процессов, лежащих в основе памяти на уровне нервной клетки. Наиболее эффективным оказался метод внутриклеточного отведения электрической активности отдельного нейрона. С его помощью можно анализировать роль синаптических процессов в изменении активности нейрона. В частности, на этой основе были установлены нейронные механизмы простой формы обучения — привыкания (см. п. 7.1.1).
Изучение нейронных основ памяти сопряжено с поиском структур, нейроны которых обнаруживают пластические изменения при обучении. Экспериментальным путем такие нейроны обнаружены у животных в гиппокампе, ретикулярной формации и некоторых зонах коры. Исследования М.Н. Ливанова и С.Р. Раевой показали, что активация оперативной памяти у человека сопровождается изменением активности нейронов многих структур мозга. При применении тестов на оперативную и непроизвольную память были обнаружены «пусковые» нейроны, расположенные в головке хвостатого ядра и передней части зрительного бугра, которые отвечали лишь на речевые команды типа: «запомните», «повторите». В контексте векторной психофизиологии (см. тему 1 п. 1.4.4) разрабатывает нейронную модель памяти Е.Н.Соколов. По его представлениям, разнообразная информация закодирована в нейронных структурах мозга в виде особыхвекторов памяти, которые создаются набором постсинаптических локусов на теле нейрона-детектора, имеющих разную электрическую проводимость. Этот вектор определяется как единица структурного кода памяти. Вектор восприятия состоит из набора постсинаптических потенциалов разнообразной амплитуды. Размерности всех векторов восприятия и всех векторов памяти одинаковы. Если узор потенциалов полностью совпадает с узором проводимостей, то это соответствует идентификации воспринимаемого сигнала.
Частотная фильтрация и память. Концепция частотной фильтрации предполагает, что обработка информации в зрительной системе осуществляется через нейронные комплексы, наделенные свойствами двумерных пространственно-частотных фильтров. Такие фильтры осуществляют анализ параметров стимула по принципу, описываемому разложением Фурье. При этом механизмы хранения энграмм находят своеобразное выражение в концепции пространственно-частотного анализа. Предполагается, что в памяти фиксируется только гармонический состав нервных импульсов, а узнавание знакомых объектов упрощается за счет того, что отношение частот внутри гармонического состава не зависит от абсолютной величины импульса. Именно поэтому для оперативной памяти требуется столь малый объем. В то же время в контексте этой модели конкретные механизмы функционирования памяти еще далеко не ясны. Однако показано, что различные пространственные частоты по-разному взаимодействуют с памятью: высокочастотная информация сохраняется в кратковременной памяти дольше, чем низкочастотная. Кроме того, нейронные механизмы, формирующие основные функциональные свойства фильтров, их пространственно-частотную избирательность, по-видимому, различным образом представлены в долговременной памяти.
Математическое моделирование памяти. Математическое моделирование на уровне суммарной биоэлектрической активности мозга применяется и к изучению памяти. Исходя из представлений об импульсном кодировании сигналов в памяти и цикличности нейронных процессов А.Н. Лебедев предлагает математическую модель, которая используя некоторые характеристики основного ритма электроэнцефалограммы — альфа-ритма — позволяет количественно оценить объем долговременной памяти и некоторые другие ее характеристики. Физиологическими основами памяти, согласно А.Н. Лебедеву, служат пачки нейронных импульсов, способные циклически повторяться. Каждая пачка импульсов — своеобразная «буква» универсального нейронного кода. Сколько разных пачек по числу импульсов в каждой, столько разных букв в нейронном коде. Пачки импульсов возникают друг за другом и образуют ограниченные цепочки. Это кодовые слова. Каждой цепочке, т.е. каждому кодовому слову, соответствует свой, порождающий его ансамбль нейронов. В результате каждому приобретенному образу памяти (слову, предмету, явлению и т.п.) соответствует свой нейронный ансамбль. Нейроны ансамбля, хранящие один образ, активизируются согласованно, циклически. Колебания клеточных потенциалов, связанные с импульсацией нейронов, создают повторяющийся узор биопотенциалов. Причем каждому образу соответствует свой собственный узор. Часть нейронов ансамбля могут «замолкать» или включаться в работу другого ансамбля, другого образа. При этом ансамбль может не только приобретать нейроны (повторение), но и терять их (забывание).
Предполагается, что работу одного ансамбля может обеспечить число нейронов от 100 до 1000. Нейроны одного ансамбля не обязательно размещаются рядом, однако часть нейронов любого образа с необходимостью располагается в ретикулярной формации ствола и промежуточного мозга, другие нейроны размещаются в старой и новой коре, в ее первичных, вторичных и третичных зонах. А.Н. Лебедев предполагает, что узоры, образованные волнами активности какого-либо ансамбля, повторяются чаще всего через 100 мс, т.е. после каждого нервного импульса клетка «отдыхает», восстанавливаясь в течении 10 мс. Это так называемая относительная рефрактерная фаза, снижающая способность нейрона включаться в коллективную деятельность под влиянием протекающих к нему импульсов от других нейронов. Таким образом синхронные импульсы многих нейронов ансамбля возникают друг за другом с промежутками около 1 мс, составляя группу, которая и является минимальной кодовой единицей памяти. Цепочка из групп, появляющаяся в одном цикле активности, может быть названа нейронным, кодовым «словом», а отдельная группа в составе слова — кодовой «буквой». Представление о циклических кодах памяти оказалось также продуктивным и для теоретического расчета быстродействия памяти, проявляющегося в скорости мнемического поиска и быстроте принятия решения в ситуации выбора (см. Хрестомат. 7.1).
7.3. Биохимические исследования памяти
Поиску специфических веществ, ответственных за хранение информации — «информационных молекул», посвящено немало исследований. Исходно эти исследования опирались на предположение, что все этапы формирования, удержания и воспроизведения энграмм можно представить в виде последовательности биохимических процессов.
«Молекулы памяти». Первые гипотезы, связывающие запечатление информации с биохимическими изменениями в нервной ткани, родились на основе широко известных в 60-е гг. опытов Г. Хидена, которые показали, что образование следов памяти сопровождается изменениями свойств РНК и белка в нейронах. Выяснилось, что раздражение нервной клетки увеличивает в ней содержание РНК и оставляет длительные биохимические следы, сообщающие клетке способность резонировать в ответ на повторные действия одних и тех же раздражителей. Таким образом, было установлено, что РНК играет важную роль в механизмах формирования и сохранения следов памяти. Однако в более поздних работах было показано, что в консолидации энграмм памяти ведущую роль играет ДНК, которая может служить хранилищем не только генетической, но и приобретенной информации, а РНК обеспечивает передачу специфического информационного кода. Высказывалось даже предположение, что неспособность зрелых нейронов делиться имеет своей целью предотвратить разрушение приобретенной информации, хранящейся в ДНК нейрона. Эти открытия имели большой научный и общественный резонанс. Некоторые исследователи, например, увлеклись идеей улучшения памяти путем введения этих биохимических компонентов в рацион питания. Однако, если иметь в виду, что крупные молекулы такого типа распадаются в кишечнике на составляющие их аминокислоты до включения их в обмен веществ потребителя, надежных результатов здесь получить было невозможно. Другим примером той же логики служили попытки переноса («транспорта памяти») от обученных животных к необученным. Методически это осуществлялась с помощью инъекций мозгового субстрата животного-донора, обученного простым навыкам, животному-реципиенту, ранее не обучавшемуся. Наибольшую популярность в связи с этим приобрели эксперименты Г. Унгара, который предпринял попытку выделить особое вещество — пептид «скотофобин», передающий информацию о страхе перед темнотой. Многочисленные проверки, последовавшие за этим открытием, не дали положительных результатов. Итак, концепции биохимического кодирования индивидуального опыта в памяти, опираются на две группы фактов: 1) образование в мозге при обучении новых биохимических факторов (например, «пептидов памяти»); 2) возможность передачи приобретенной информации необученному мозгу с помощью этих факторов. Однако идея существования биохимических факторов, способных к сохранению и переносу информации, большинством исследователей воспринимается критически. В настоящее время считается, что гипотеза молекулярного кодирования индивидуального опыта не имеет прямых фактических доказательств. Несмотря на то, что установлена существенная роль нуклеиновых кислот и белков в механизмах научения и памяти, предполагается, что принимающие участие в формировании новой ассоциативной связи РНК и белки специфичны лишь по отношению к функциональному изменению участвующих в процессе синапсов и неспецифичны по отношению к самой информации.
Медиаторные системы. Медиаторам — химическим посредникам в синаптической передаче информации — придается большое значение в обеспечении механизмов долговременной памяти. Основные медиаторные системы головного мозга — холинэргическая и моноаминоэргическая (включает норадреноэргическую, дофаминэргическую и серотонинэргическую) — принимают самое непосредственное участие в обучении и формировании энграмм памяти. Так, экспериментально установлено, что уменьшение количества норадреналина замедляет обучение, вызывает амнезию и нарушает извлечение следов из памяти. Р.И. Кругликов (1986) разработал концепцию, в соответствии с которой в основе долговременной памяти лежат сложные структурнохимические преобразования на системном и клеточном уровнях головного мозга. При этом холинэргическая система мозга обеспечивает информационную составляющую процесса обучения. Моноаминоэргические системы мозга в большей степени связаны с обеспечением подкрепляющих и мотивационных составляющих процессов обучения и памяти. Показано, что под влиянием обучения увеличивается количество холинорецепторов, т.е. рецепторов, расположенных на теле нейрона и отвечающих за обнаружение медиатора ацетилхолина. В процессе образования условного рефлекса повышается чувствительность соответствующих нейронов к ацетилхолин у, что облегчает обучение, ускоряет запоминание и способствует более быстрому извлечению следа из памяти. В то же время вещества, препятствующие действию ацетилхолина, нарушают обучение и воспроизведение, вызывая амнезию (потерю памяти).
Важно подчеркнуть, что холинэргическая система испытывает на себе модулирующее влияние со стороны моноамионоэргической системы. Под действием этих влияний может изменяться активность холинэргических синапсов и запускаться цепь биохимических внутриклеточных процессов, приводящих к более эффективному образованию энграмм.
Значение биохимических исследований памяти. Биохимические методы, позволяющие проникнуть в последовательность процессов, разыгрывающихся в синаптических мембранах с последующим синтезом новых белков, привлекают многих исследователей памяти. На этом пути ожидаются новые яркие открытия. Предполагается, например, что для различных видов памяти в ближайшем будущем будут выявлены различия в биохимических процессах. Тем не менее следует подчеркнуть, что интенсивные биохимические исследования привели к явной переоценке и автономизации клеточно-молекулярного уровня изучения механизмов памяти. Как указывает С. Роуз, эксперименты, проводимые только на клеточном уровне, слишком ограничены, и, по-видимому, не способны ответить на вопрос — как мозг человека запоминает, например, сложные симфонические партитуры, или извлекает из памяти данные, необходимые для разгадывания простого кроссворда (см. Хрестомат. 7.3). Для более полного знания о специфике функционирования процессов памяти необходим переход на уровень сложных мозговых систем, где многие нейроны соединены между собой морфологическими и функциональными связями. При этом психофизиологические исследования на здоровых людях позволяют изучать процессы переработки и хранения информации, а изучение больных с различного рода амнезиями, возникающими после повреждения мозга, позволяют глубже проникать в тайны памяти. Память нельзя рассматривать как нечто статичное, находящееся строго в одном месте или в небольшой группе клеток. Память существует в динамичной и относительно распределенной форме. При этом мозг действует какфункциональная система, насыщенная разнообразными связями, которые лежат в основе регуляции процессов памяти.
Нейрональная пластичность Многие ученые считают, что обучение и память основаны исключительно на синаптической пластичности (потенциации и депрессии передачи сигнала в синапсах).
Нейроны, при этом, выполняют лишь интегративную функцию для возбуждающих и тормозных синаптических токов, результатом которой является вероятность возникновения потенциала действия в аксоне. Это является значительным упрощением, как и представление о том, что тоническая активация внесинаптических рецепторов лишь меняет возбудимость нейронов посредством шунтирующего тока. Изменение биофизических свойств (проводимости, возбудимости, потенциала, ионных градиентов) мембраны нейрона или его отдельных компартментов (сомы, ответвлений дендритов) при активации внесинаптических рецепторов диффузный нейропередатчиком может изменять характеристики входящих синаптических токов (подавление, усиление, укорачивание, удлинение).
Причем эти изменения происходят благодаря свойствам мембраны нейронов лежащей за пределами синапсов. Так, можно представить, что в условиях неизменной синаптической передачи, синаптические токи могут быть подавлены или усилены в зависимости от компартмента нейрона. Процесс изменения синаптического тока в зависимости от состояния мембраны нейрона-мишени можно терминологически определить как «нейрональная обработка синаптического сигнала». Одним из способов нейрональной обработки синаптического сигнала является вышеупомянутый ГАМКергический тонический ток. В своей недавней работе Митчелл и Сильвер (S. J. Mitchell and R. A. Silver) показали, что этот ток определяет усиление выходного сигнала нейрона (частоту нейрональных разрядов) и регулирует обработку частотно-кодированной синаптической информации. Активация внесинаптических рецепторов является свойством не только ГАМКергической системы. Во многих лабораториях мира, в том числе и нашей было показано, что возбуждающий нейропередатчик — глутамат, может покидать синаптическую щель и активировать внесинаптические рецепторы. Если нейрональная обработка синаптического сигнала будет изменять свои параметры во времени, то этот процесс будет называться «нейрональной пластичностью». Такие изменения могут носить как кратковременный, так и долговременный характер. Примером, долговременного изменения нейрональной обработки сигнала является появление тонического ГАМКергического тока при эпилепсии в нейронах, которых он отсутствовал в нормальном состоянии (см. выше).
Это указывает на роль нейрональной пластичности в патологических состояниях мозга. Тем не менее, не существует принципиального противоречия в утверждении, что нейрональная пластичность, аналогично синаптической (а возможно вместе с синаптической), вовлекается в процессы обучения и памяти. 6. Нейрональная пластичность и возбудимость нейрона На первый взгляд может показаться, что возбудимость нейрона и нейрональная пластичность по сути одно и тоже. Нейрон получает синаптический сигнал и далее генерирует или не генерирует спайк (потенциал действия).
Возбудимость нейрона обычно оценивают по параметру ВПСП (возбуждающий постсинаптический потенциал) — спайк передачи. Если вероятность генерации спайка на одинаковый по амплитуде ВПСП повышается, то это называется ВПСП-спайк потенциацией. Если вероятность снижается, то это называется ВПСП-спайк депрессией. Нейрональная пластичность процесс более сложный. Она, в частности, определяет с каких компартментов нейрона информацию следует усилить, а с каких подавить. Это позволят нейрону выбирать входящие сигнальные пути, что не обязательно отразится на его возбудимости, но может определить, какой информационный поток в локальной нейрональной сети получит приоритет. Важно отметить, что в отличие от синаптической, нейрональная пластичность не специфична для отдельных синапсов, а специфична для целого нейрона или его компартментов (например, может шунтировать одновременно тормозные и возбуждающие синаптические токи).
Нейрональная пластичность является типом обработки сигнала в мозге, который существует параллельно с пластичностью синапса и пластичностью ВПСП-спайк передачи (возбудимости нейрона).
Не следует забывать, при этом, что все классификации условны. В реальном нейроне эти типы обработки сигнала тесно взаимодействуют друг с другом. Например, изменение электрохимических градиентов потенциал-образующих ионов может привести к изменению возбудимости нейрона, поскольку изменится порог генерации Ca2+ спайков в дендритах и/или Na+/K+ спайков в аксоне. С другой стороны, изменение градиентов будет изменять электродвижущую силу для ионов, принимающих участие в синаптических событиях. Так, низкое содержание ионов хлора внутри нейрона и высокое снаружи делает синаптические токи, опосредованные рецепторами ГАМК, тормозными (гиперполяризующими).
Низкая концентрация хлора внутри клетки достигается за счет хлорного насоса — белкового комплекса, расположенного в нейрональной мембране. Если этот комплекс перестает работать, то градиент хлора выравнивается, а величина ГАМКергического торможения снижается. При этом возникает вероятность ситуации, что ГАМК может быть даже возбуждающим (деполяризующим) нейропередатчиком. Было показано, что ГАМК является исключительно возбуждающим нейропередатчиком на ранних стадиях развития мозга и становится преимущественно тормозным во взрослом состоянии.
Привыкание (психология)
[править | править вики-текст]
Материал из Википедии — свободной энциклопедии
Привыкание (габитуация) это постепенное уменьшение ответной реакции как результат продолжающейся или повторяющейся стимуляции в нормальных условиях (не сопровождается подкреплением стимула).
Привыкание свойственно всем живым существам. Следует отличать привыкание от процессов мышечного утомления, а также от сенсорной адаптации. Последняя представляет собой процессы происходящие с сенсорными органами, тогда как привыкание это психические процессы выработки ответной реакции на стимул. Привыкание в отличие от сенсорной адаптации может осуществляться как сознательно так и неосознанно. Привыкание сходно с процессами затухания выработанной ответной реакции. Выработанная ответная реакция может проявиться даже после прекращения действия стимула. Если стимуляция слабая то привыкание происходит быстрее. Сильная стимуляция может и не сопровождаться привыканием. При прочих равных условиях стимуляция с более высокой частотой повторения вызывает более быстрое привыкание.
Определяющим привыкание параметрами является частота и интенсивность стимула. В то же время привыкание происходит при условии соразмерности частоты поведению.
Противоположностью привыканию является отвыкание, когда изменившийся сходный стимул вызывает ответную реакцию. Исследуя процессы отвыкания связанные с проявлением сильного измененного стимула Томпсон и Спенсер обнаружили, что отвыкание фактически не является в полном смысле отвыканием так как не прерывает процесс привыкания. В этом случае происходит наложение независимого процесса повышенной возбудимости то есть сенсибилизация.[1]
Нейробиология
Согласно Соколову Е.Н. повторение стимула приводит к развитию нейронной модели стимула в структурах переднего мозга. Если стимул связан с такой моделью тогда не проявляется ответная реакция о чём могут свидетельствовать показания электроэнцефалографии отмечающие отсутствие изменений активности мозга. Если же стимул или изменения стимула не связаны с выработанной нейронной моделью, то отраженные изменения стимула усиливается через ретикулярные формации(ретикулярная активирующая система) среднего мозга что приводит более сильной реакции.[2] Процессы формирования нейронной модели можно свести к процессамсинаптической депрессии (См. синаптическая пластичность) в цикле стимуляции нейронной активности.
Процессы привыкания связаны как с сенсорной так и с моторной системами мозга. Последние исследования показывают, что привыкание к стимулам сенсорной системы мозга происходит до привыкания моторной системы. Ретикулярные ядра выполняют функцию моста между сенсорной и моторной системами.[3]. В исследованиях аплизии Кэндэл (Kandel) показали, что привыкание является пресинаптическим процессом связанным с уменьшением вероятности высвобождениянейромедиаторов. Последнее является следствием повторяющейся активации высвобождения нейромедиаторов.
Исследования связи привыкания и сенсибилизации в зрительном внимании у детей показали, что привыкание чаще всего происходит в случаи простых стимулов. Сложные стимулы провоцируют как привыкание так и сенсибилизацию.[4]
Сенситизация — повышение чувствительности, повышение нормы эмоционального реагирования, когда нервная система начинает реагировать на небольшой сигнал существенно активнее, чем обычно.
Как форма научения, обычно такая реакция происходит после неожиданного крайне сильного сигнала (обычно негативного) или в тревожной обстановке.
Например, если вы идете домой днем, когда светло и много народу — вас не испугает даже подошедший близко незнакомый человек. А вот если вы идете затемно, да еще и одни, у вас состояние другое (у вас сенситизация) — и ваша реакция будет совсем другой.
Клеточные аналоги условного рефлекса
Выделяют две основные гипотезы обучения: синоптическую, предполагающую, что механизмом формирования условных рефлексов является изменение эффективности синапсов, и мембранную, утверждающую, что основным механизмом условного рефлекса является изменение свойств возбудимой постсинаптической мембраны. Как мы увидим далее, такое деление весьма условно, однако представляется удобным для первого знакомства с проблемой.
Нервные механизмы обучения обеспечиваются благодаря двум основным свойствам нервной системы – реактивности и пластичности. Если реактивность – это способность отвечать на раздражения, то пластичность – это способность изменять реактивность под влиянием последовательных раздражений. В свою очередь, реактивность определяется возбудимостью постсинаптической мембраны и эффективностью соответствующих синапсов. Возбудимость нейрона измеряется силой тока, необходимой для генерации потенциала действия, а синоптическая эффективность – амплитудой моносинаптического постсинаптического потенциала.
Уже давно высказывалось предположение о том, что в основе обучения лежит изменение эффективности межклеточных контактов, или проторение новых контактов. На рис. 77 приведены основные схемы. Простейшим является синапс Дж. Экклса (а), повышающий свою эффективность после собственной активации (фасилитация).
Такие синапсы могут служить для объяснения феноменов посттетанической потенциации. Дополнением к ним являются синапсы с постепенным снижением эффективности при повторной активации (б).
Они могут обеспечивать эффекты габитуации и дегабитуации. Как мы уже отмечали, при выработке условных рефлексов необходимо временное совпадение пресинаптической активации с активацией подкрепляющих систем, которые могут оказывать также пресинаптическое действие (в).
Д.О. Хебб (1969) главным условием синаптической модификации считал совпадение пресинаптической активности с разрядом постсинаптического нейрона (г).
Затем к этому была добавлена модель Альбуса, связанная не с увеличением эффективности синапса, а с ее снижением (д).
Наконец, в качестве синтетической схемы (е) рассматриваются случаи изменений синаптической эффективности с участием трех факторов – сенсорного притока, активации подкрепляющей системы и достижения результата.
Вопрос о характере преобразований нейрональной активности в динамике выработки условного рефлекса начал исследоваться в связи с внедрением в практику физиологического эксперимента тонких методов микроэлектродного анализа и последующей компьютерной обработки импульсных реакций нейронов. Попытки использовать внутриклеточную регистрацию постсинаптических электрических потенциалов привели к необходимости проводить исследования либо на относительно простых нервных сетях, либо на центральных нейронах высших млекопитающих с максимальным упрощением требуемых для анализа межнейронных отношений. Такие упрощенные модели принято рассматривать в качестве клеточных аналогов условного рефлекса.
Группой венгерских физиологов (О. Фехер, А. Барани, 1981) разработана модель ассоциативного обучения, при котором сигнал меняет свое действие под влиянием последующего безусловного эффекта.
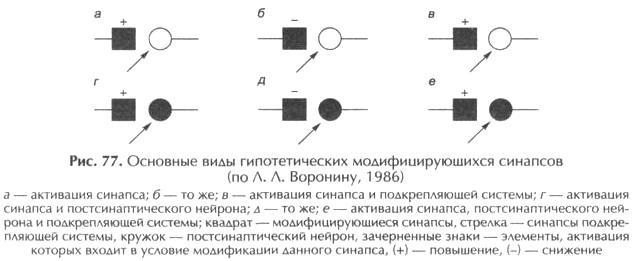
Рассмотрим процедуру обусловливания при “условном” раздражении вентролатерального ядра таламуса (ВЛ) и антидромной стимуляции пирамидного тракта (“безусловный стимул”).
Сначала, как показано на рис. 78, электрическая стимуляция ВЛ с частотой 1 с сопровождается только подпороговыми ПСП. Последующее антидромное раздражение пирамидного тракта вызывает один потенциал действия. По мере сочетаний раздражения таламуса с антидромной стимуляцией амплитуда и/или длительность ТПСП увеличиваются и остаются таковыми в течение нескольких минут. Амплитуда ВПСП изменяется также, но к 50-му сочетанию ВПСП достигает порога генерации и “условный” сигнал начинает вызывать 1-2 потенциала действия. Эффект антидромного раздражения при этом не меняется. К 100-му сочетанию амплитуда ВПСП еще больше возрастает, а число потенциалов действия достигает трех. Если проводить угашение, то есть подавать одно “условное” раздражение, не сопровождая его антидромной стимуляцией, то амплитуда ВПСП и число генерируемых потенциалов действия уменьшается, и спустя 350 изолированных раздражений таламического ядра ответная “условная” реакция исчезает. Внутриклеточная поляризация исследуемого нейрона, приводившая к генерации потенциалов действия, усиливала эффект раздражения вентролатерального таламуса. Однако ни многократное применение только “условного” стимула, ни псевдообусловливание (процедура применения условного и безусловного раздражителей в случайном порядке) не привели к генерации потенциалов действия исследуемого нейрона. Уровень мембранного потенциала и сопротивление мембраны при выработке временной связи менялись по-разному в различных клетках. Это позволило думать об определяющей роли при выработке временной связи избирательных синоптических сдвигов.
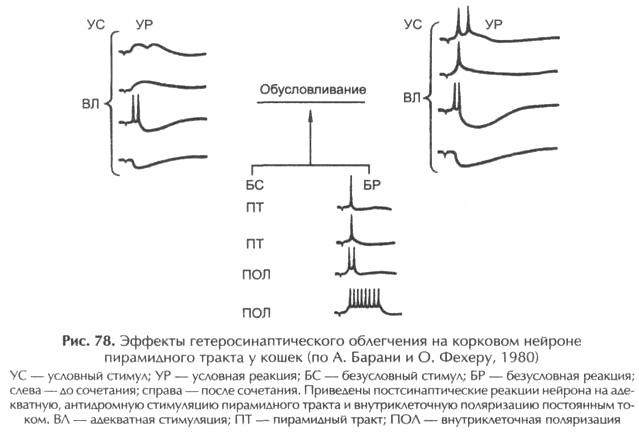
Было показано, что положительные и дифференцировочные связи с исследуемым нейроном опосредуются разными синапсами независимо друг от друга. Е.И. Соколов (1931) справедливо полагает, что элементарным локусом пластического изменения является не нейрон, а синапс. Речь идет о синапсе Хебба, который селективно изменяется, если за синаптической посылкой следует спайковый разряд нейрона. Такое явление получило наименование гетеросинаптического облегчения.
Этот же феномен использовался для объяснения ряда эффектов обусловливания в другой модели (Л.Л. Воронин, 1982).
Синаптические аналоги условного рефлекса создавались в зоне коркового представительства передней конечности бодрствующего кролика сочетаниями электрического раздражения двух точек коры. Условным стимулом служило подпороговое (неэффективное) раздражение коры на некотором расстоянии от исследуемого коркового нейрона, а безусловным – пороговое для постсинаптической реакции раздражение поверхности коры над исследуемым корковым нейроном. При активирующих формах условной связи в ответ на условный стимул возникали ВПСП, амплитуда которых постепенно возрастала до критического уровня генерации потенциалов действия (рис. 79).
При отмене подкрепления, то есть при угашении, амплитуда ВПСП постепенно уменьшалась с прекращением генерации спайковых разрядов.

При тормозных формах временной связи условный стимул постепенно вызывал развивающееся угнетение механизма генерации импульсных разрядов, которое не сопровождалось существенной гиперполяризацией мембраны клетки или достоверным уменьшением ВПСП. При этом установлено, что дифференцирование одним и тем же нейроном подкрепляемых и неподкрепляемых стимулов реализуется различными синаптическими входами и поддерживается благодаря механизмам гетеросинаптического облегчения.
Одной из моделей аналога временной связи на микропопуляции нейронов коры больших полушарий является микроионофоретическая аппликация медиаторов (глутамата и ацетилхолина), используемых в качестве аналогов ассоциируемых раздражителей (Б.И. Котляр, 1986).
Установлено, что отдельные нейроны микропопуляции в радиусе действия медиаторов обнаруживают специфические ассоциативные перестройки своей активности. Последние могут заключаться в следующем: изменения реактивности к сигнальному раздражителю, воспроизведение реакции на подкрепляющий раздражитель в ответ на действие условного сигнала и реакция на время в границах действия ассоциируемых раздражителей.
В.С. Русинов (1969) пришел к выводу, что суммационный рефлекс и доминанта представляют собой начальные стадии образования любого условного рефлекса и сами по себе могут служить удобной моделью для изучения простых форм временной связи. Им была разработана модель корковой поляризационной доминанты, состоящая в том, что поляризация постоянным током участков сенсомоторной коры кролика при действии ранее неэффективных раздражителей – световых вспышек, приводила теперь к ритмическому сокращению передней конечности (корковая проекция которой поляризовалась) в такт световой стимуляции. Установившаяся под влиянием поляризации новая форма реагирования на раздражитель сохраняется до 30 мин после выключения тока.
Эта модель послужила существенным толчком для выявления пластических характеристик корковых нейронов. Установлено, что поляризация слабым постоянным током увеличивает активность полисенсорных нейронов, что является одним из непременных механизмов возникновения движения на ранее индифферентный раздражитель. Причем функциональные и морфологические сдвиги происходят не только в пирамидных нейронах, но и в интернейронах и глиальных клетках, которые также подвергаются поляризации.
Все основные свойства доминанты – повышенная возбудимость, стойкость возбуждения, способность к суммированию возбуждений, инерции, торможению и растормаживанию – были обнаружены на вышеописанной модели. В доминантном очаге происходит установка на определенный уровень стационарного возбуждения, способствующий суммированию ранее подпороговых возбуждений и переводу на оптимальный для данных условий ритм работы, когда этот очаг становится наиболее отзывчивым и тормозятся другие работающие очаги. Иными словами, образуется единый функционально организованный ансамбль, или система. Подчеркивается особая роль нейронов нижних слоев коры, которые дольше сохраняют ритм раздражения, чем клетки верхних слоев. Обращается серьезное внимание на существование в системе базальных дендритов пирамидных клеток дендродендритических контактов, которые рассматриваются как важнейший субстрат межнейронных отношений. В.С. Русинов полагает, что наиболее адекватным отражением функционального состояния системы нейронов является уровень их постоянного потенциала, а следовательно, электротонические влияния имеют наиболее существенное значение для образования временной связи. Поляризационный очаг, созданный постоянным током, служит удачной моделью для изучения поляризационно-электротонической гипотезы формирования доминанты и простых форм временных связей (В.С. Русинов, 1969).
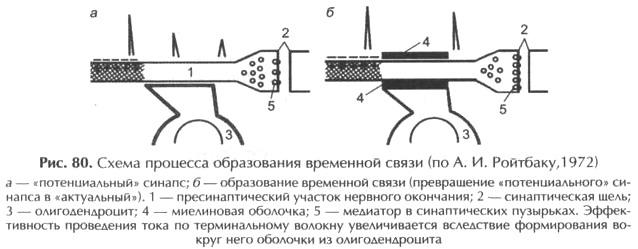
Специальное внимание к нейроглиальным взаимоотношениям при формировании клеточных аналогов условного рефлекса привлек А.И. Ройтбак (1969).
Он выдвинулглиальную гипотезу, согласно которой в основе образования и укрепления временных связей лежит миелинизация центральных аксонов, возбуждающихся от условного раздражения. При действии безусловного раздражения в соответствующих областях коры происходит деполяризация глиальных клеток: возбуждение большого числа нервных элементов импульсами из специфических и неспецифических путей приводит к выделению в межклеточные щели и аккумуляции ионов калия. Автор полагает, что деполяризация олигодендроцитов служит сигналом к образованию миелина; условием миелинизации является совпадение во времени деполяризации олигодендроцитов и физико-химических изменений в прилегающих немиелинизированных пресинаптических терминалях после их предшествующего возбуждения (рис. 80).
В последние годы широко развернулись исследования механизмов временных связей на животных с относительно простой нервной системой. Наиболее удобными для экспериментирования оказались моллюски. Простейшей моделью служил полностью изолированный нейрон висцерального ганглия виноградной улитки. Локальная аппликация ацетилхолина сочеталась с внутриклеточной инъекцией деполяризующего тока, в результате было показано, что изолированные нейроны моллюсков (то есть лишенные своих сетевых свойств) не способны формировать ассоциативный ответ в виде воспроизведения безусловной реакции сигнальным раздражителем.
Клеточные механизмы образования условного рефлекса на морском моллюске аплизии изучались Э. Кенделом (1980).
Исследовался оборонительный рефлекс: условный сигнал – умеренной силы раздражение сифона, вызывающее втягивание жабры в мантийную полость, безусловный раздражитель – сильная стимуляция хвоста. В результате сочетаний защитная реакция на стимуляцию сифона усиливалась. Удобство этой модели состояло в том, что был точно определен нейронный состав рефлекторной дуги, в которую входят 24 сенсорных нейрона, образующие прямые связи с интернейронами и 6 моторными нейронами, иннервирующими жабру и сифон. Стимуляция хвоста возбуждает нейроны, контактирующие с пресинаптическими окончаниями сенсорных нейронов.
В данной модели обнаружились явления сенситизации, привыкания и нейрогуморального (субклеточно-молекулярного) воздействия безусловного раздражителя на всех этапах выработки временной связи. Суммация заключалась в неспецифическом гетеросинаптическом облегчающем эффекте – увеличении ВПСП, связанном с увеличением выброса медиатора из пресинаптического окончания. Суммация этих процессов приводит к возникновению длительной деполяризации, составляющей сущность проторения путей при образовании временной связи. Процессы привыкания связаны с ограничением потоков кальция в пресинаптические окончания, что приводит к гиперполяризации нейронов и снижению их ответа на безусловный раздражитель. Нейрогуморальный эффект безусловного раздражителя достигался благодаря механизмам регуляторных нейропептидов (см. гл. 9).
Э. Кендел (1980) считает, что в основе условной реакции втягивания жабры у аплизии лежит механизм пресинаптического облегчения (сенситизации), который формируется при конвергенции раздражителей, приводящих к спайковой активности нейрона, и активации этого же нейрона через модулирующий вход от безусловного раздражителя. Аналогичный механизм допускается и у позвоночных животных, хотя прямые доказательства этого отсутствуют.
Какими бы информативными для задач общей нейрофизиологии ни были аналитические подходы к исследованию наркотизированных или обездвиженных препаратов позвоночных и беспозвоночных животных с использованием различных моделей и аналогов условных рефлексов, их конечной целью является расшифровка тех феноменов мультинейронной активности, которые регистрируются на бодрствующем мозге при выполнении животным того или иного целостного условно-рефлекторного акта. Сегодня мы еще не можем использовать весь арсенал сведений аналитической нейрофизиологии для понимания клеточной активности бодрствующего мозга. Различия двух основных подходов базируются на степени проявления свойства пластичности, а именно – высокой изменчивости реакций одного и того же нейрона у бодрствующего животного и значительно большей стабильности реагирования нервных клеток у наркотизированных препаратов.
Научение
Понятия “память” и “научение” психологи и нейробиологи традиционно относят к поведенческим категориям. Они применимы для характеристики целостного организма. Память и научение – неотделимые процессы. Научение обеспечивает постоянное пополнение и изменение наших знаний, а также приобретение новых навыков, умений. В отличие от научения процесы памяти ответственны не только за усвоение (фиксацию), но и за сохранение и воспроизведение (извлечение) информации. В самом широком смысле слова научение можно определить как приспособительное изменение поведения, обусловленное прошлым опытом. Память необходима для научения, так как она представляет собой механизм, с помощью которого накапливается прошлый опыт, который может стать источником адаптивных изменений поведения.
Научение – совокупность процессов, обеспечивающих приобретение индивидуальной (фенотипической) памяти, вызывающей приспособительную модификацию поведения.
Научение требует определенного времени, условий и реализуется с помощью нейрофизиологических механизмов разного уровня (межклеточного, внутриклеточного, молекулярного).
6.3.1. Виды научения
Существует много разновидностей научения. Они могут быть разделены на несколько групп. Наиболее часто выделяют простое научение, к которому относят привыкание, сенситизацию, ассоциативное научение, включающее выработку классического условного рефлекса, инструментального (или оперантного) рефлекса, одномоментное обучение (на аверсивном подкреплении), и сложное научение (импринтинг, латентное обучение, обучение на основе подражания, когнитивное обучение: формирование декларативной памяти).
Привыкание как простейшая форма научения выражается в ослаблении поведенческой реакции при повторных предъявлениях стимула. От утомления и истощения привыкание отличается тем, что реакцию вновь можно вызвать простым изменением стимула.
Э. Кэндел и его сотрудники исследовали привыкание оборонительной реакции у аплизии. Защитная реакция морского моллюска в виде втягивания сифона и жабры вызывалась тактильным раздражением этих органов струей холодной воды. Многократное применение такой стимуляции уменьшало величину рефлекторной реакции втягивания. Эффект ослабления реакции мог быть кратковременным, но при достаточно большом числе повторений снижение реактивности оборонительной реакции могло сохраняться в течение нескольких недель (долговременное привыкание).
Ярким примером поведенческого привыкания является угасание безусловного ориентировочного рефлекса (или его отдельных компонентов).
С повторением стимула теряется новизна, что и приводит к привыканию. Привыкание в системе ориентировочного рефлекса получило название негативного научения, состоящего в том, что стимул по мере его повторения теряет способность вызывать ту реакцию, которую он ранее вызывал. Привыкание или угасание ориентировочного рефлекса связывают с формированием “нервной модели стимула” – его многомерной энграммы, которая и тормозит систему активации ориентировочного рефлекса (см. главу “Внимание”).
Сенситизация – другая форма простейшего научения, выражающаяся в усилении рефлекторной реакции под влиянием сильного или повреждающего постороннего стимула. Сенситизация – это не просто противоположность привыкания. Она является результатом активации модулирующей системы мозга, возникшей на сильный побочный раздражитель. Усиление рефлекса вызвано изменением функционального состояния организма.
6.3.2. Нейронные феномены пластичности
Пластичность – фундаментальное свойство клетки, которое проявляется в относительно устойчивых модификациях реакций нейрона и во внутриклеточных его преобразованиях, обеспечивающих изменение эффективности и направленности межнейронных связей.
Свойство пластичности нейрона лежит в основе процессов научения и памяти целостного организма, проявляющихся на поведенческом уровне. Выделяют несколько основных феноменов пластичности: привыкание, сенситизацию, клеточные аналоги ассоциативного обучения, явления долговременной потенциации и долговременной депрессии, пластичность пейсмекерного механизма клетки.
Привыкание нейрона выражается в постепенном ослаблении его реакции на повторяющийся раздражитель. Восстановление реакции происходит в результате изменения стимула или применения нового, а также после прекращения привычной стимуляции. Время восстановления реакции (секунды – недели) зависит от стимуляции и изучаемого объекта.
Сенситизация нейрона – временное усиление его реакции или появление ее на ранее неэффективный стимул, возникающее в результате какого-либо сильного воздействия (например, электрического тока). Время ее сохранения – от нескольких секунд до дней и недель. Сенситизацию связывают с активацией модулирующих нейронов, вызываемой сильным биологически значимым воздействием.
Долговременная потенциация (long-term-potentiation, LTP) впервые описана для нейронов гиппокампа. В 1973 г. Т. Блис и Т. Лемо, работая в лаборатории П. Андерсена в Осло (Норвегия), впервые показали, что нейроны гиппокампа обладают выраженными пластическими свойствами, нужными для обучения. Применив высокочастотную электрическую стимуляцию, чтобы вызвать короткую серию ПД в пресинаптическом нейроне, они получили эффект усиления ответов постсинаптического нейрона на последующие одиночные импульсы, приходящие от пресинаптической клетки. Эффект мог сохраняться в течение часов-недель. Они назвали такое усиление синаптической проводимости долговременной потенциа-цией, предположив, что постсинаптическая клетка выделяет ретроградный мессенджер,который способен проникать через мембрану пресинаптической клетки. Этим мессенджером, возможно, является окись азота, которая определяет пресинаптическое облегчение у гип-покампальных нейронов млекопитающих.
Долговременная потенциация (ДВП) в гиппокампе удовлетворяет выдвинутому Д. Хеббом критерию обучения – совпадению пресинаптической и постсинаптической активности нейрона. Этот принцип был сформулирован Д. Хеббом в 1949 г. в книге “Организация поведения” в качестве условия, необходимого для образования новых нейронных ансамблей в процессе обучения.
В опытах на срезах гиппокампа П. Андерсен и его сотрудники (Andersen P. et al., 1977) показали, что долговременная потенциа-ция обусловлена усиленным выделением медиатора из пресинап-тических окончаний. С помощью электронного микроскопа была подтверждена связь долговременной потенциации с увеличением шипиков на дендритах нейронов гиппокампа и изменением структуры синапсов.
Позже ДВП была обнаружена в других структурах мозга – миндалине, мозжечке. ДВП относится к определенному синаптичес-кому контакту. Она выявлена у идентифицированного синапса в простой нервной системе моллюска.
Долговременную потенциацию (ДВП) можно обнаружить по увеличению амплитуды и крутизны ВПСП, повышению вероятности генерации ПД, а также по увеличению числа спайков на один тестирующий стимул. Долговременная потенциация часто используется как удобная модель для изучения механизмов научения.
Долговременная депрессия (long-term-depression, LTD) состоит в длительном снижении проводимости через синапс. Сравнительно недавно долговременная депрессия (ДВД) обнаружена в коре мозжечка у клеток Пуркинье. Ее можно вызвать совместной низкочастотной стимуляцией (1-4 Гц) двух входов к клеткам Пуркинье: афферентного входа через параллельные волокна к ее дендри-там и входа от лазящих волокон из нижней оливы, посылающей сигналы о совершаемых двигательных рефлексах.
Сигнал от лазящего волокна увеличивает приток ионов Са2+ к клеткам Пуркинье. Это вызывает в нейроне каскад преобразований, где главная роль принадлежит процессам фосфорилирования-дефос-форилирования. Предполагают, что ДВД возникает вследствие того, что ионы Са2+ активируют белок фосфатазу, который обеспечивает дефосфорилирование рецепторов на дендритах клеток Пуркинье, снижая тем самым их синаптическую проводимость. Не исключается возможность и второго механизма торможения клеток Пуркинье через долговременную потенциацию тормозного входа к ним от корзин-чатых интернейронов. ДВД возникает только на тех синапсах параллельных волокон к клеткам Пуркинье, возбуждение которых по времени совпадает с сигналом, приходящим из нижней оливы.
Таким образом, долговременная депрессия в клетках Пуркинье – тот механизм пластических изменений в нейроне, благодаря которому в мозжечке формируются условные рефлексы.
Клеточные аналоги ассоциативного обучения можно получить во многих структурах мозга. Так, 40% нейронов гиппокампа способны к обучению (Котляр Б.И., 1986).
Процедура формирования условного рефлекса у нейрона требует, чтобы к нему последовательно, с небольшой задержкой приходили сигналы от условного и безусловного стимулов. Условным стимулом может быть любое сенсорное или электрическое раздражение, которое вызывает лишь подпороговую реакцию (ВПСП).
Безусловный раздражитель должен вызывать у нейрона спайковые разряды (ПД).
Под влиянием сочетания “индифферентного” раздражителя с безусловным нейрон, способный к ассоциативному обучению, начинает отвечать на условный стимул реакцией, которая ранее возникала только на безусловное раздражение.
У отдельного нейрона можно выработать не только классический условный рефлекс, но и инструментальный. Впервые это показали исследователи из Мичигана Джеймс и Мариана Олдс в 1961 г., а затем и другие исследователи. В опытах Э. Фетца и М.Э. Бейкер (Fetz E., Baker M., 1973) с помощью вживленных электродов регистрировали активность нейронов в двигательной коре у бодрствующей обезьяны. Когда частота ПД случайно превышала некоторый уровень, шкала прибора перед обезьяной освещалась и животное получало положительное подкрепление – несколько капель фруктового сока. В результате такой процедуры активность нейрона возрастала до того уровня, который подкреплялся. С отменой подкрепления частота ПД возвращалась к исходному, фоновому значению (угасание инструментального рефлекса).
Параллельная регистрация ЭМГ и механограммы мышцы задней стопы показала, что возрастание частоты спайков нейрона двигательной коры, вызванное обучением, несколько опережает начало электрической и механической активности мышцы. Это позволило утверждать, что инструментальный условный рефлекс был выработан у нейрона, который и запускал двигательную реакцию, а не наоборот.
Многие исследования были посвящены формированию аналогов условного рефлекса на различных модельных системах: срезах мозга, изолированной нервной системе моллюска, синапти-чески связанных нейронах. Особенно впечатляющими оказались результаты изучения клеточных механизмов пластичности на полностью изолированной соме нейрона. Этот препарат впервые был получен у аплизии и применен Р. Баумгартеном с соавторами (Baumgarten R.I. et al., 1971).
На виноградной улитке ассоциативное обучение у полностью изолированного нейрона было исследовано Т.Н. Греченко (1979).
Сому нейрона, изолированную от отростков, получают при помощи обработки ганглиев виноградной улитки протеомитическим ферментом – трипсином с последующим выдерживанием в термостате при температуре 37-38°С в течение 15-20 мин. После такой процедуры нейроны лишаются отростков. Изолированный нейрон может состоять только из сомы или из сомы с культей аксона. Он сохраняет присущую ему фоновую ативность, тип хемочувствительности.
В качестве условного стимула (УС) использовались микроаппликация медиаторов (ацетилхолина или серотонина) или прямая деполяризация током через внутриклеточный электрод. Все условные стимулы вызывали лишь ВПСП и были подпороговыми для ПД. В качестве безусловного стимула (БС) применялся раздражитель большой силы, который вызывал устойчивую генерацию ПД. Интервал времени между УС и БС 5-500 мс, интервал между парами стимулов – от 40 с до 3 мин. Через 10-15 сочетаний условный стимул начинал вызывать реакцию, характерную для БС. Условный ответ у разных нейронов сохранялся в течение 5-40 мин. Дополнительное обучение увеличивало время сохранения следового эффекта. Неподкрепление вело к постепенному угасанию условной реакции. 80% нейронов (из 503) продемонстрировали феномен отсроченного обучения. Условный ответ после прекращения процедуры обучения появлялся не сразу, а спустя некоторое время. Интервал от момента прекращения обучения до достижения максимального следового эффекта для разных нейронов составлял от 1-3 до 60 мин и более (Греченко Т.Н., Соколов Е.Н., 1987).
Был выявлен локальный характер обучения на одном нейроне. Условные и безусловные раздражители действуют через отдельные локусы мембраны, и условный ответ формируется только между теми участками мембраны, к которым направлены УС и БС. Поэтому на одном изолированном нейроне можно выработать параллельно несколько различных условных рефлексов, включая диф-ференцировку, а также получить их селективное угасание. Эти результаты невозможно объяснить механизмом пресинаптической пластичности: полностью изолированные нейроны лишены каких-либо контактов с другими нейронами. Они указывают на роль пост-синаптического и, вероятно, молекулярных механизмов в становлении следов памяти.
6.3.3. Пластичность пейсмекерного механизма
Особый вид пластичности обнаруживают пейсмекерные нейроны, способные к генерации эндогенного ритма.
Пейсмекеры (водители ритма) представляют собой источники активности мозга. Различают пейсмекеры в виде нейронной сети и в виде отдельных нейронов, способных генерировать ритмическую спайковую активность.
Самый простой сетевой пейсмекер – это по крайней мере два нейрона, соединенных обратной положительной связью. Такая система обеспечивает межклеточную реверберацию сигнала (его многократное отражение).
Реверберация реализуется через нервную сеть, так как сигнал выходит за пределы мембраны нейрона, где он первично возник. Воздействуя на один из нейронов сетевого пейсмекера, можно управлять его работой, включая или выключая реверберацию сигнала.
Другой тип пейсмекера состоит из одного нейрона, в котором возникает внутриклеточная реверберация импульса. Ее появление обусловлено двусторонним электротоническим распространением ПД вдоль аксона к его окончанию и в обратном направлении к соме. Достигая аксонного холмика, он вызывает генерацию нового ПД. Так возникает серия импульсов за счет челночного движения ПД. Но так как при распространении ПД вдоль аксона он активирует ионные каналы Са2+, то в результате увеличения притока ионов кальция внутрь клетки открываются кальций-зависимые К-каналы. Ионы К выходят в экстраклеточное пространство, и возникает волна гиперполяризации, которая ограничивает длительность ПД и ВПСП. Если заблокировать калиевые каналы, то клетка перейдет в режим реверберации. Таким образом, Са2+-зависимые К-каналы представляют собой звено, через которое можно включать и выключать режим внутриклеточной реверберации возбуждения.
Третий тип пейсмекера – это нейрон, в сому которого встроен генератор ритмической активности. Такие нейроны получили название пейсмекерных нейронов.Потенциалы, которые они генерируют (синусоидальные колебания на частоте около 0,1-10 Гц), относятся к эндогенным и могут быть выявлены у полностью изолированной сомы (Греченко Т.Н., Соколов Е.Н., 1987).
Пейсме-керные потенциалы модулируют уровень мембранного потенциала. В отсутствие внешнего источника возбуждения синусоидальные эндогенные потенциалы периодически достигают порога генерации ПД, что и определяет появление у них ритмических спайко-вых разрядов. Предполагают, что пейсмекерная активность таких нейронов связана с колебаниями электрогенного активного транспорта ионов. Активный транспорт восстанавливает нарушенный после генерации ПД градиент ионов Na и К, имеющийся по обе стороны мембраны, и тем самым предохраняет нейрон от перевозбуждения. Пейсмекерная активность такого нейрона отражает ритм открывания и закрывания Са2+-каналов, которые взаимодействуют с Са2+-зависимыми К-каналами.
К факторам, запускающим генерацию эндогенного ритма, относятся синаптические и гуморальные влияния, действие пептидов, диффузное внесинаптическое влияние медиаторов. Они включают и выключают режим пейсмекерной активности. Управление пейсмекерной активностью экстраклеточными факторами обеспечивается взаимодействием локуса генерации эндогенного ритма с хемовозбудимой и электровозбудимой мембраной. Показана зависимость ритмической активности нейронного пейсмекера от уровня деполяризации нейрона. При слабой деполяризации основной вклад в генерацию эндогенных потенциалов вносят низкопороговые Са2+-каналы, при высоких значениях деполяризации – высокопороговые. С заменой низкопороговых Са2+-каналов на высокопороговые частота эндогенного ритма возрастает.
Пейсмекерные нейроны используются для управления реакциями организма. С их помощью реализуются генетические программы (например, локомоции).
Благодаря их зависимости от си-наптических влияний они могут обеспечивать более гибкое применение генетических программ в адаптивном поведении.
Пейсмекерный механизм обладает пластичностью, которая выражается в том, что ответ нейрона меняется по мере повторения действия одиночных раздражителей. В том случае, если ответ пейсмекерного нейрона слабеет от применения к применению, этот процесс по аналогии с угасанием поведенческой реакции называют привыканием. Кроме привыкания, имеет место обратный процесс возрастания реакции нейрона от применения к применению. Его называют фасилитацией. Пластические реакции у пейсмекерного нейрона могут обеспечиваться изменением как возбудимости пейсмекерного механизма, так и эффективности синоптической передачи. Пластические свойства пейсмекерного механизма существенно расширяют адаптационные возможности организма в манипуляции генетическими программами поведения.
|
Долговременная потенциация Долговременная потенциация (синоним: длительная потенциация; англ. Long—term potentiation, сокр.LTP) — усиление синаптической передачи между двумя нейронами, сохраняющееся на протяжениидлительного времени после воздействия на синаптический проводящий путь. LTP участвует в механизмахсинаптической пластичности, обеспечивающих нервную систему живого организма возможностьюадаптироваться к изменяющимся условиям внешней среды. Большинство теоретиков нейрофизиологииполагают, что долговременная потенциация совместно с долговременной депрессией лежат в основеклеточных механизмов памяти и обучения. Долговременная потенциация была открыта норвежским ученым Терье Лёмо (Terje Lømo) в 1966 году припроведении опытов на гиппокампах кроликов[1]. Первая научная работа, описывающая процесс LTP, былаопубликована Терье Лёмо и Тимоти Блиссом (Timothy Bliss) в 1973 году В шестидесятых годах в нейрофизиологической литературе стали появляться отдельные сообщения о том, что многократная, производимая с достаточной частотой стимуляция нервных путей к некоторым областям коры головного мозга приводит к длительному усилению спонтанной электрической активности таких областей. Это явление — в сущности, повышение эффективности передачи между пре- и постсинаптическими клетками — получило название потенциации. Не может ли такая потенциация быть одной из форм нейрофизиологической памяти? В 1973 году в очень часто цитируемой теперь статье Тим Блисс из Национального института медицинских исследований в Лондоне и Терье Лёмо описали совместную работу, проведенную ими в лаборатории Пера Андерсена в Осло. Они обнажали у наркотизированных кроликов гиппокамп и идущие к нему нервные пути и подводили к одному из этих путей — «перфорантному» (perforant) — стимулирующие электроды, а регистрирующие электроды вводили в ту область гиппокампа, где перфорантный путь образует синапсы (в зубчатую извилину, рис. 9.4). Когда они после этого стимулировали перфорантный путь залпом электрических импульсов частотой 10-100 в секунду и длительностью до 10 секунд, наблюдалось необычайно продолжительное (до 10 часов) усиление активности нейронов зубчатой извилины гиппокампа [15]. Авторы назвали этот феномен долговременной потенциацией (сокращенно ДВП). На самом деле ДВП длится гораздо дольше десяти часов; этот эффект наблюдали и у наркотизированных животных с вживленными электродами, причем в этом случае потенциация сохранялась иногда на протяжении 16 недель после первоначальной кратковременной стимуляции. По-видимому, краткая электрическая стимуляция гиппокампа приводила к изменению электрических свойств его клеток. Блисс, Лемо, Андерсен и многие другие нейробиологи сразу же заинтересовались этим явлением. Это был сильно выраженный эффект — специфический, воспроизводимый и, сверх того, поддававшийся физиологическому, а позднее также биохимическому, фармакологическому и морфологическому исследованию. Гиппокамп млекопитающих был уже хорошо известной структурой, его нервные связи, входные и выходные нервные пути подробно картированы и легко распознаваемы на различных препаратах, хотя индивидуальные нейроны не поддавались такой прямой идентификации, как у аплизии. Продолжительное изменение выходной клеточной реакции на определенное входное воздействие служит по меньшей мере ярким примером нервной пластичности; больше того, весьма специфичная форма, которую приобретает реакция, может рассматриваться как проявление памяти. Было уже достаточно хорошо известно, что и у человека, и у других млекопитающих гиппокамп имеет какое-то отношение к памяти. Не может ли долговременная потенциация (ДВП) служить механизмом образования энграмм? Не могут ли физиологи использовать ее хотя бы как модель для изучения памяти? Блисс и Лемо явно склонялись к этому, считая свою процедуру стимуляции аналогом «условного раздражения»; однако они заключили свою статью загадочной оговоркой: «Другое дело — использует ли интактное животное в реальной жизни то свойство, которое было выявлено путем синхронной повторной стимуляции группы нервных волокон, нормальная активность которых неизвестна» [15]. ДВП легко вызывать и изучать классическими методами нейрофизиологии, поэтому вряд ли стоит удивляться ее популярности в качестве потенциальной модели памяти. В ближайшие годы после первых наблюдений Блисс в Лондоне, Андерсен в Осло и все большее число исследователей в других лабораториях стали в мельчайших подробностях изучать физиологию ДВП. Было показано, что этот феномен выявляется не только у наркотизированных и ненаркотизированных кроликов, крыс и других лабораторных животных, но и в препаратах in vitro. Гиппокамп легко извлечь из мозга вместе с его входными нервными путями, в том числе и перфорантным путем. Благодаря трехмерной структуре гиппокампа можно получать тонкие срезы с интактными путями к клеткам того же среза, как показано на рис. 9.4. Такие срезы культивировали in vitro и изучали их электрические свойства. Так забытая с пятидесятых годов методика Мак-Илвейна снова вошла в моду у нейробиологов в конце семидесятых годов. Надлежащая стимуляция нервных путей в таких срезах тоже приводила к ДВП, которая сохранялась до отмирания среза. Характеристики ДВП примерно одинаковы и в срезах, и в интактном мозгу. Во-первых, в обоих случаях реакция специфична для определенных путей. Иными словами, она развивается только в тех клетках, к которым подводится «условный раздражитель», а не распространяется от клетки к клетке, т. е. это результат функционирования сети специфических связей, а не волна диффузной активности. Такую специфичность можно изящно продемонстрировать благодаря наличию нескольких различных подводящих путей к отдельным участкам даже в одном срезе гиппокампа. Во-вторых, для индукции ДВП требуется повторное воздействие достаточно частых импульсов: то же число импульсов, поступающих с меньшей частотой, не вызывает эффекта. Таким образом, существует частотный порог индукции ДВП. При частоте импульсов выше пороговой ДВП может развиваться либо постепенно, либо по принципу «все или ничего» в зависимости от ритма, интенсивности и частоты условных стимулов. Развитие ДВП происходит по меньшей мере в две (возможно, в три) фазы, с коротким периодом инициации и более длительной фазой устойчивой реакции, которые рассматриваются как аналоги перехода от кратковременной памяти к долговременной. Третий и, пожалуй, самый интересный момент с точки зрения клеточной аналогии памяти состоит в следующем. В начале восьмидесятых годов было показано, что существует ассоциативная форма ДВП. При этом слабая стимуляция, недостаточная сама по себе для поддержания ДВП, может стать активирующим фактором, если сочетается с сильной стимуляцией через второй путь [16]. Оба вида стимуляции должны совпадать или ассоциироваться во времени, как условный и безусловный раздражители при ассоциативном научении. Можно даже получить такую форму ассоциативного научения, в которой смешаны поведенческие и нейрофизиологические стимулы. Крыса может научиться переходить границу между двумя отделениями ящика в ответ на сигнал, состоящий только из залпа импульсов на гиппокамп, в качестве безусловного раздражителя [17]. Все эти особенности ДВП, взятые вместе, вызывали повышенный интерес к ее изучению по меньшей мере как одной из интригующих моделей памяти. |
Пластичность синапсов
В ходе функционирования синапсы подвергаются функциональным и морфологическим перестройкам. Этот процесс назван синаптической пластичностью. Наиболее ярко такие изменения проявляются при высокочастотной, или тетанической активности, являющейся естественным условием функционирования синапсов in vivo. Например, частота импульсации вставочных нейронов в ЦНС достигает 1000 Гц. Пластичность (рис. 6-10) может проявляться либо в увеличении (облегчении, потенциации), либо уменьшении (депрессии) эффективности синаптической передачи. Выделяют кратковременные (длятся секунды и минуты) и долговременные (длятся часы, месяцы, годы) формы синаптической пластичности. Последние интересны тем, что они имеют отношение к процессам научения и памяти.
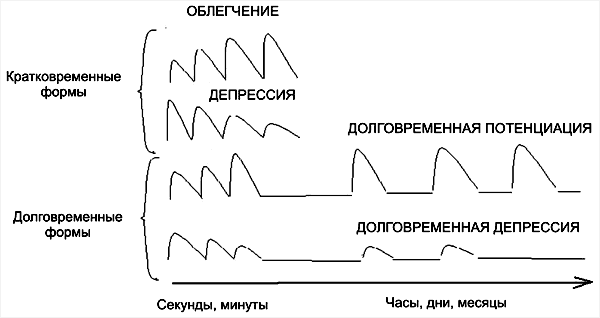
Рис. 6–10. Формы синаптической пластичности
Кратковременные формы синаптической пластичности
К ним относятся облегчение, потенциация, депрессия и привыкание.
Облегчение. В процессе активности в синапсах с исходно низким уровнем секреции нередко происходит увеличение амплитуды постсинаптического потенциала (ПСП).
Этот процесс — облегчение — имеет пресинаптическую природу и объясняется теорией «остаточного кальция». Согласно этой теории, в процессе высокочастотной активности в пресинаптической терминали наблюдается повышение концентрации Са2+, вследствие чего происходит увеличение вероятности освобождения квантов нейромедиатора.
Потенциация, посттетаническая потенциация (сенситизация). Увеличение ПСП при высокочастотной активности может иметь и постсинаптическую природу. Такой вид пластичности связан с повышением чувствительности постсинаптических рецепторов к нейромедиатору и называется потенциацией. Величина ПСП может некоторое время (секунды и минуты) оставаться повышенной и после окончания тетанической активности. Это посттетаническая потенциация (в ЦНС — сенситизация).
Депрессия и привыкание (габитуация). В синапсах с исходно высоким уровнем секреции высокочастотная активность может приводить к уменьшению величины ПСП. Этот процесс — депрессия — связан преимущественно с истощением запаса нейромедиатора в пресинаптическом нервном окончании. Депрессия является одним из механизмов привыкания (габитуации).
Долговременные формы синаптической пластичности
Долговременная потенциация — быстро развивающееся устойчивое усиление синаптической передачи в ответ на высокочастотное раздражение. Этот вид пластичности может продолжаться дни и месяцы (рис. 6–11).
Долговременная потенциация наблюдается во всех отделах ЦНС, но наиболее полно изучена на глутаматергических синапсах в гиппокампе.
Существует три основных подтипа ионотропных глутаматных рецепторов: NMDA (чувствительны к N-метил-D-аспартату), AMPA (связываются с ?-амино-3-гидрокси-5-изоксазолпропионовой кислотой) и каинатные рецепторы. NMDA- и AMPA–рецепторы играют ключевую роль в возникновении и проявлении долговременной потенциации.
Механизм долговременной потенциации
— При высокочастотной стимуляции нейронов гиппокампа выделяется большое количество глутамата, деполяризуется постсинаптическая мембрана и происходит активация NMDA–рецепторов. Значительный кальциевый ток через эти каналы приводит к повышению концентрации ионов Са2+ в постсинаптическом нейроне.
— Ионы Са2+ связываются с внутриклеточным белком — кальмодулином (Кальмодулин — Ca2+-связывающий белок; связывание с Ca2+ в цитоплазме клеток изменяет его конформацию и превращает его в активатор ферментов, например, фосфодиэстераз или киназы лёгкой цепи миозина в ГМК; регулятор процесса сокращения ГМК и многих внутриклеточных событий).
Образовавшийся комплекс активирует фермент — Са2+-кальмодулинзависимую протеинкиназу II.
— Са2+-кальмодулинзависимая протеинкиназа II фосфорилирует AMPA–рецепторы. После фосфорилирования возрастает их ионная проводимость, что приводит к увеличению постсинаптического ответа на каждый квант нейромедиатора. Кроме того, данный фермент мобилизует дополнительные AMPA–рецепторы из цитоплазмы в постсинаптическую мембрану, что приводит к увеличению квантового состава ПСП.
Долговременная депрессия также возникает в ответ на высокочастотное раздражение и проявляется в виде длительного ослабления синаптической передачи. Этот вид пластичности имеет сходный механизм с долговременной потенциацией, но развивается при более низкой внутриклеточной концентрации ионов Са2+.
В заключение приведём данные о продолжительности различных событий, происходящих в синапсах на их постинаптической стороне: из рис. 6–11 видно, что продолжительность процессов имеет широкий разброс — от 1 мсек (деполяризация постсинаптической мембраны за счёт ионотропных рецепторов) до дней (модуляция синаптической передачи).
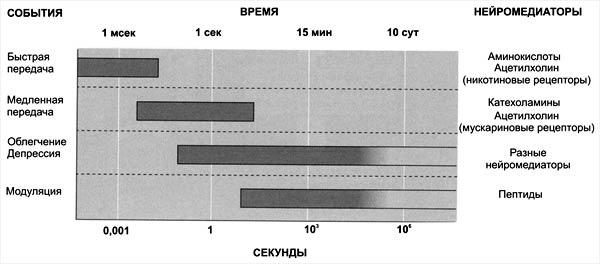
Рис. 6–11. Сравнительная продолжительность различных событий в синапсах [2]. Логарифмическая шкала.